
У каких растений в ксилеме отсутствуют сосуды
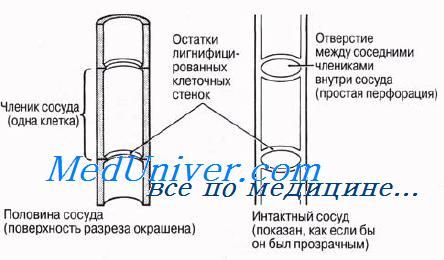
Ксилема. Строение ксилемы. Функции ксилемы.Ксилема выполняет в растении две основные функции: по ней движется вода вместе с растворенными минеральными веществами и она служит опорой органам растения. Таким образом, ксилема играет в растении двоякую роль — физиологическую и структурную. В состав ксилемы входят гистологические элементы четырех типов: трахеиды, сосуды, паренхимные клетки и волокна. На рис. 6.9 эти гистологические элементы представлены и поперечном и продольном разрезах. Трахеиды ксилемыТрахеиды — это одиночные лигнифицированные клетки веретеновидной формы. Концы соприкасающихся трахеид перекрываются так же, как и заостренные концы волокон склеренхимы. Это придает трахеидам механическую прочность и обеспечивает органам растения опору. Трахеиды — мертвые клетки; в зрелом состоянии их просвет ничем не заполнен. Среди водопроводя-ших клеток сосудистых растений трахеиды представляют первичную примитивную форму; у древних сосудистых растений это единственные водопроводящие клетки. Из них развились описанные ниже сосуды и волокна ксилемы высших растений. Несмотря на свой примитивный характер, трахеиды, несомненно, функционируют эффективно; об этом свидетельствует тот факт, что у голосеменных растений доставка воды от корней к надземным частям обеспечивается исключительно трахеидами, а ведь большинство голосеменных — древесные породы. Вода движется по пустым просветам трахеид, не встречая на своем пути помех в виде живого содержимого. Из одной трахеиды в другую она переходит либо через поры, через их «замыкающие пленки», либо через нелигнифицированные части клеточных стенок. Характер лигнификации (одревеснения) клеточных стенок трахеид близок к тому, который описан ниже для сосудов. На рисунке представлено строение трахеид. У покрытосеменных число трахеид по сравнению с числом сосудов относительно невелико. Сосуды считаются более эффективным приспособлением для транспорта воды, нежели трахеиды; появление сосудов связано, как полагают, с тем, что у покрытосеменных с их большой листовой поверхностью транспира-ция идет более активно. Сосуды ксилемыСосуды — характерные проводящие элементы ксилемы покрытосеменных. Они представляют собой очень длинные трубки, образовавшиеся в результате слияния ряда клеток, соединившихся «конец в конец». Каждая из клеток, образующих сосуд ксилемы, соответствует трахеиде и называется члеником сосуда. Однако членики сосуда короче и шире трахеид. Первая ксилема, появляющаяся в растении в процессе сто развития, носит название первичной ксилемы; она закладывается у кончика корня и на верхушке побегов. Дифференцированные членики сосудов ксилемы появляются рядами на концах прокамбиальных тяжей. Сосуд возникает, когда соседние членики в данном ряду сливаются в результате разрушения перегородок между ними. Внутри сосуда сохраняются в виде ободков остатки разрушенных торцевых стенок. Слияние члеников сосудов изображено на рисунке.
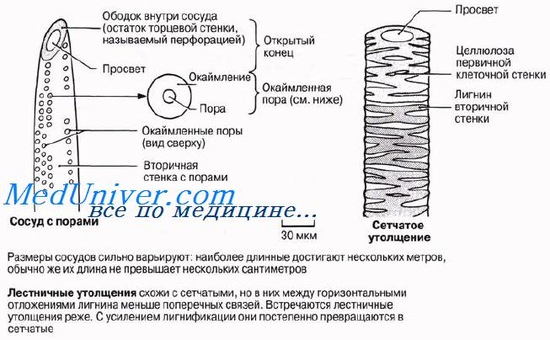
Протоксилема и метаксилемаПервые по времени образования сосуды — протоксилема — закладываются на верхушке осевых органов, непосредственно под верхушечной меристемой, там, где окружающие их клетки еше продолжают вытягиваться. Зрелые сосуды про-токсилемы способны растягиваться одновременно с вытягиванием окружающих клеток, поскольку их целлюлозные стенки еще не сплошь одревеснели —лигнин откладывается в них лишь кольцами или по спирали (рис. 6.12). Эти отложения лигнина позволяют трубкам сохранять достаточную прочность во время роста стебля или корня. С ростом органа появляются новые сосуды ксилемы, которые претерпевают более интенсивную лигнификацию и завершают свое развитие в зрелых частях органа; так формируется ме-гаксшема. Тем временем самые первые сосуды протоксилемы растягиваются, а затем разрушаются. Зрелые сосуды метаксилемы не способны растягиваться и расти. Это мертвые, жесткие? полностью одревесневшие трубки. Если бы их развитие завершалось до того, как закончилось вытягивание окружающих живых клеток, то они бы очень сильно мешали этому процессу. У сосудов метаксилемы обнаруживаются три главных типа утолщений: лестничные, сетчатые и точечные. Длинные полые трубки ксилемы — идеальная система для проведения воды на большие расстояния с минимальными помехами. Так же как и в трахеидах, вода может переходить из сосуда в сосуд через поры или через неодревеснев-шие части клеточной стенки. Вследствие одревеснения клеточные стенки сосудов обладают высокой прочностью на разрыв, что тоже очень важно, потому что благодаря этому трубки не спадаются, когда вода движется в них под натяжением (разд. 13.4). Вторую свою функцию — механическую — ксилема выполняет также благодаря тому, что она состоит из ряда одревесневших трубок. В первичном теле растения ксилема в корнях занимает центральное положение, помогая корню противостоять тянущему усилию надземных частей, изгибающихся под порывами ветра, В стебле проводящие пучки либо образуют по периферии кольцо, как у двудольных, либо располагаются беспорядочно, как у однодольных; в обоих случаях стебель пронизывается отдельными тяжами ксилемы, обеспечивающими ему определенную опору. Особенно важное значение опорная функция ксилемы приобретает там, где имеет место вторичный рост. Во время этого процесса быстро нарастает количество вторичной ксилемы; к ней переходит от колленхимы и склеренхимы роль главной механической ткани, и именно она служит опорой у крупных древесных и кустарниковых пород. Рост стволов в толщину определяется в известной мере нагрузками, которым подвергается растение, так что иногда наблюдается дополнительный рост, смысл которого состоит в усилении структуры и обеспечении ей максимальной опоры.
Древесинная паренхима ксилемыДревесинная паренхима ксилемы содержится как в первичной, так и во вторичной ксилеме, однако в последней ее количество больше и роль важнее. Клетки древесинной паренхимы, подобно любым другим паренхимным клеткам, имеют тонкие целлюлозные стенки и живое содержимое. Во вторичной ксилеме имеются две системы паренхимы. Обе они возникают из меристемати-ческих клеток, называемых в одном случае лучевыми инициалями, а вдругом — веретеновидны-ми инициалями (гл. 22). Лучевая паренхима более обильна. Она образует радиальные слои ткани, так называемые сердцевинные лучи, которые, пронизывая сердцевину, служат живой связью между сердцевиной и корой. Здесь запасаются различные питательные вещества, скапливаются таннины, кристаллы и т. п., и здесь же осуществляется радиальный транспорт питательных веществ и воды, а также газообмен по межклетникам. Из веретеновидных инициалей обычно развиваются сосуды ксилемы и ситовидные трубки флоэмы вместе с их клетками-спутницами, однако время от времени они дают начало также и паренхимным клеткам. Эти паренхимные клетки образуют во вторичной ксилеме вертикальные ряды. Древесинные волокна ксилемыПолагают, что древесинные волокна, так же как и сосуды ксилемы, ведут свое происхождение от трахеид. Они короче и уже трахеид, а стенки их гораздо толще, но поры их сходны с порами, имеющимися в трахеидах, и на срезах волокна иногда трудно отличить от трахеид, поскольку между теми и другими есть ряд переходных форм. Древесинные волокна очень напоминают уже описанные волокна склеренхимы; их торцевые стенки также перекрываются. В отличие от сосудов ксилемы древесинные волокна не проводят воду; поэтому у них могут быть гораздо более толстые стенки и более узкие просветы, а значит, они отличаются и большей прочностью, т. е. придают ксилеме дополнительную механическую прочность. – Также рекомендуем “Флоэма. Строение флоэмы. Функции флоэмы.” Оглавление темы “Ткани. Питание клетки.”: |
Источник
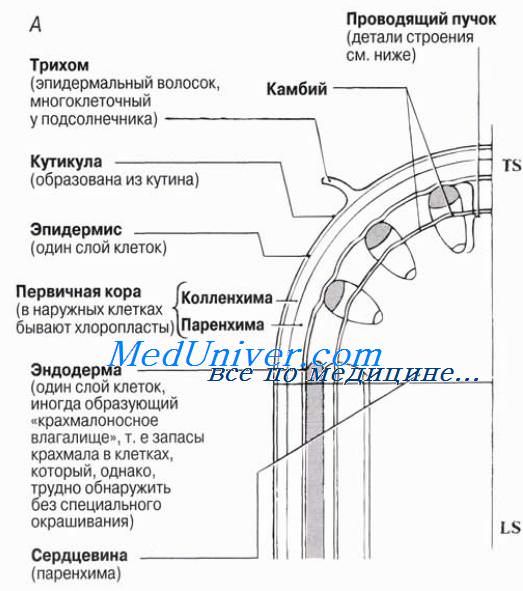
Подъем воды по ксилеме растений.Ксилема цветковых растений содержит два типа проводящих воду структур — трахеиды и сосуды. В статье мы уже говорили о том, как выглядят эти структуры в световом микроскопе, а также на микрофотографиях, полученных с помощью сканирующего электронного микроскопа. Строение вторичной ксилемы (древесины) рассматривается в статье. Ксилема и флоэма образуют проводящую ткань высших, или сосудистых, растений. Эта ткань состоит из так называемых проводящих пучков, строение и распределение которых в стеблях двудольных растений с первичным строением показано на рисунке. То, что вода поднимается именно по ксилеме, нетрудно продемонстрировать, погрузив побег срезанным концом в разбавленный водный раствор красителя, например эозина. Подкрашенная жидкость, распространившись вверх по стеблю, заполнит сеть пронизывающих листья жилок. Если затем сделать тонкие срезы и рассмотреть их в световом микроскопе, то окажется, что краситель находится в ксилеме. Более эффектное доказательство подъема воды по ксилеме дают опыты с «кольцеванием». Такие опыты проводили задолго до того, как стали применяться радиоактивные изотопы, позволяющие очень легко проследить путь веществ в живом организме. В одном из вариантов опыта с одревесневшего стебля снимают узкое кольцо коры вместе с лубом, т. е. флоэмой. Довольно долго после этого находящиеся выше вырезанного кольца побеги продолжают расти нормально: следовательно, такое кольцевание не влияет на подъем воды по стеблю. Однако, если, приподняв лоскут коры, вырезать из-под него сегмент древесины, т. е. ксилемы, то растение быстро завянет. Таким образом, вода движется в побеги из почвы именно по этой проводящей ткани. Любая теория, объясняющая транспорт воды по ксилеме, не может не учитывать следующие наблюдения.
1. Анатомические элементы ксилемы — тонкие мертвые трубки, диаметр которых варьирует от 0,01 мм в «летней» древесине до 0,2 мм в «весенней» древесине. Удовлетворительное объяснение этим фактам дает теория сцепления (когезии), или теория натяжения. Согласно этой теории, подъем воды от корней обусловлен ее испарением клетками листа. Как мы уже говорили в статье, испарение снижает водный потенциал клеток мезофилла, прилежащих к ксилеме, и вода поступает в эти клетки из ксилемного сока, водный потенциал которого выше; при этом она проходит через влажные клеточные стенки у концов жилок, как показано на рисунке. Ксилемные сосуды заполняет сплошной столб воды; по мере того как вода выходит из сосудов, в этом столбе создается натяжение; оно передается вниз по стеблю до самого корня благодаря сцеплению (когезии) молекул воды. Эти молекулы стремятся «прилипнуть» друг к другу, потому что они полярны и притягиваются друг к другу электрическими силами, а затем удерживаются вместе водородными связями. Кроме того, они притягиваются к стенкам ксилемных сосудов, т. е. происходит их адгезия (прилипание) к ним. Сильная когезия молекул воды означает, что ее столб трудно разорвать — у него высокий предел прочности при растяжении. Растягивающее напряжение в клетках ксилемы приводит к генерированию силы, способной сдвигать весь водяной столб вверх по механизму объемного потока. Снизу вода поступает в ксилему из соседних клеток корня. При этом очень важно, что стенки ксилемных элементов жесткие и не спадаются при падении давления внутри, как это бывает, когда сосешь коктейль через мягкую соломинку. Жесткость стенок обеспечивается лигнином. Доказательством того, что жидкость внутри ксилемных сосудов сильно напряжена (растянута), служат суточные колебания диаметра древесных стволов, измеряемые инструментом под названием дендрограф. Минимальный диаметр отмечен днем, когда интенсивность транспирации наивысшая. Натяжение столба воды в ксилемном сосуде немного втягивает внутрь его стенки (из-за адгезии), и сочетание этих микроскопических сжатий дает фиксируемую прибором общую «усадку» ствола. Оценки прочности на разрыв столба ксилемного сока варьировали от 3000 доЗО 000 к Па, причем более низкие значения получены позднее. В листьях зарегистрирован водный потенциал порядка —4000 кПа, и прочность столба ксилемного сока, вероятно, достаточна, чтобы выдержать создающееся натяжение. Не исключено, конечно, что столб воды может иногда разрываться, особенно в сосудах большого диаметра. Критики изложенной теории подчеркивают, что любое нарушение непрерывности столба сока должно немедленно останавливать весь поток, так как сосуд заполнится воздухом и паром (явление кавитации). Кавитацию может вызвать сильное сотрясение, изгибание ствола, а также дефицит воды. Хорошо известно, что на протяжении лета содержание воды в стволе дерева постепенно снижается, древесина заполняется воздухом. Этим пользуются лесозаготовители, потому что такие деревья легче сплавлять. Однако разрыв водного столба в части сосудов слабо влияет на общую скорость объемного потока. Возможно, дело в том, что вода перетекает в параллельно проходящие сосуды или же обходит воздушную пробку, продвигаясь по соседним паренхимным клеткам и по стенкам. Кроме того, согласно расчетам, для поддержания наблюдаемой скорости потока вполне достаточно, чтобы в каждый момент времени функционировала хотя бы небольшая доля ксилемных элементов. У некоторых деревьев и кустарников вода перемещается лишь по более молодой наружной древесине, называемой заболонью. У дуба и ясеня, например, проводящую функцию выполняют в основном сосуды текущего года, а остальная часть заболони играет роль водного резерва. Новые ксилемные сосуды образуются на протяжении всего вегетационного периода, но главным образом в его начале, когда скорость водного потока максимальна. Вторая сила, обеспечивающая движение воды по ксилеме, — корневое давление. Его можно обнаружить и измерить в тот момент, когда срезают крону, а штамб с корнями некоторое время продолжает выделять сок из сосудов ксилемы. Этот процесс подавляется ингибиторами дыхания, например цианидом, и прекращается при недостатке кислорода и понижении температуры. Работа такого механизма, по-видимому, обусловлена активной секрецией солей и других водорастворимых веществ в ксилемный сок. В результате его водный потенциал падает, и вода поступает в ксилему из соседних клеток корня путем осмоса. Этот механизм создает гидростатическое давление порядка 100-200 кПа (в исключительных случаях 800 кПа); одного его для подъема воды по ксилеме обычно недостаточно, однако у многих растений оно, несомненно, способствует поддержанию ксилемного тока. У медленно транспирирующих травянистых форм этого давления вполне хватает, чтобы вызвать у них мутацию. Так называется выделение воды на поверхности растения1 в виде капель жидкости, а не пара. Все условия, тормозящие транспира-цию, например слабая освещенность и высокая влажность, способствуют гуттации. Она обычна у многих видов дождевых тропических лесов и часто наблюдается на кончиках листьев у всходов трав. – Также рекомендуем “Поглощение воды корнями. Апопластный транспорт в корне.” Оглавление темы “Транспорт у растений.”: |
Источник
Основным элементом ксилемы у высшего двудольного растения является сосуд.
Как и у ситовидных трубок, составляющие сосуд клетки-членики расположены наподобие звеньев цепи вдоль длинной оси органа, соединяясь друг с другом своеобразно модифицированными поперечными стенками. Членики сосудов во многих случаях также вытянуты, но не менее часто бывают и короткими.
Поперечные перегородки между члениками сосуда к тому времени, когда сосуд окончательно сформируется, продырявливаются, и протопласты клеток, формирующих сосуд, разрушаются. Если поперечные стенки члеников сосуда разрушаются полностью, образуется одно крупное отверстие, называемое простой перфорацией. Известны также и множественные перфорации, образующиеся в том случае, если оболочка не разрушается, а продырявливается во многих местах. Отверстия при множественной перфорации разбросаны в беспорядке (сетчатая перфорация) или располагаются правильными рядами (лестничная перфорация).
Утолщения на стенках сосудов могут быть весьма различного характера. Наиболее просто устроены спиральные утолщения, а также родственные им кольчатые утолщения. И те и другие свойственны наиболее рано возникающим анатомическим элементам ксилемы у высших растений. Древние формы растений с определенно выраженной ксилемой также имеют ксилемные элементы со спирально-кольчатыми утолщениями. От сосудов со спиральными утолщениями ряд постепенных переходов ведет к сосудам с округлыми окаймленными порами. На продольных разрезах через ксилему осевого органа какого-либо двудольного можно найти почти все градации утолщений. На рисунке при рассмотрении справа налево видны два крайних сосуда со спиральными утолщениями (4), причем более тонкий из них имеет несколько сильнее растянутые завороты спирали. Следующий сосуд (3) содержит так называемые лестничные утолщения. Сосуд (2) имеет вполне оформленные окаймленные поры— округлые и многочисленные. Это — пористый сосуд. Иногда переход от сосудов со спиральными утолщениями к пористым сосудам бывает очень резок (например, в стебле льна), чем нарушается онтогенетическая последовательность образования утолщений. Тип утолщений в значительной мере зависит от того, какой анатомический элемент находится по соседству с сосудом.
На рисунке изображен сосуд, к которому с одной стороны (3) примыкает паренхима, а с другой — либриформ (2), у которого вообще образуется мало пор.
Там, где к сосуду примыкает либриформ, на стенках сосуда пор совсем не образуется, а со стороны паренхимы стенки сосуда усеяны порами. Еще значительнее усложняется характер утолщений на стенках сосудов при соприкосновении их с клетками сердцевинных лучей, в свою очередь, достаточно разнообразных по очертаниям и структуре.
Кроме сосудов с продырявленными поперечными перегородками (перфорациями), в ксилеме большинства растений существуют другие водопроводящие элементы — трахеиды. Каждая трахеида — это отдельная мертвая прозенхимная клетка с более или менее заостренными концами. В отличие от сосудов поперечные стенки трахеид не разрушаются и перфораций не возникает. Утолщения стенок трахеид совершенно такие же, как и стенок сосудов. Среди трахеид встречаются клетки и со спиральными утолщениями, и с округлыми окаймленными порами. Трахеиды в некоторой мере обладают способностью к скользящему росту и могут врастать или между другими анатомическими элементами, или между другими трахеидами. Скользящим ростом обладают лишь молодые формирующиеся анатомические элементы.
Между трахеидами и либриформом существует ряд переходных форм. У семенных растений трахеиды в большем или меньшем количестве в зависимости от вида перемежаются с сосудами и другими элементами ксилемы. У папоротников, плаунов, хвощей и голосеменных растений трахеиды составляют почти всю массу ксилемы.
Как сосуды, так и трахеиды служат для проведения воды по растению в различных направлениях. Вместе с водой проводятся различные растворимые в воде минеральные и органические вещества. Наравне с различными солями, поглощаемыми корнями из почвы, по водоносным элементам, особенно по сосудам, могут передвигаться и растворы сахаров. Например, весной, в период так называемого весеннего «плача» растений, пасока, содержащая, кроме других веществ, и сахара, продвигается в значительной мере по сосудам.
Подобно тому, как у ситовидных трубок имеются спутники, к сосудам часто примыкают паренхимные клетки, образующие обкладку сосуда. В зависимости от общей структуры ксилемы обкладка состоит или из одних паренхимных клеток, или в нее входят еще либриформ, трахеиды и более мелкие сосуды. Паренхима не только окружает сосуды, но и рассеяна среди ксилемных элементов; в последнем случае она называется древесинной паренхимой. Клетки ее имеют одревесневшие оболочки с простыми порами, но со стороны сосуда им соответствует окаймленная пора, которая сочетается с каждой простой порой. Протопласт клеток древесинной паренхимы очень долго не разрушается. Клетки паренхимы в ксилеме служат местом отложения запасов, так же как и паренхима, входящая в состав других тканей.
Иногда и на оболочках древесинной паренхимы развиваются окаймленные поры. Такие клетки не сохраняют протопласт в жизненном состоянии, так как служат передаточными пунктами в продвижении воды к соответствующим тканям.
Клетки паренхимы, непосредственно примыкающие к сосуду, находятся в тесном контакте с растворами, заполняющими сосуды, и в зависимости от условий то конденсируют в своих пластидах поступающие к ним углеводы и прочие вещества, то отдают эти вещества в полость сосуда. Специальными наблюдениями установлено, что в клетках паренхимы, примыкающих к сосуду и сообщающихся с ним через односторонние окаймленные поры, крахмал, сахар и прочие пластические вещества в течение года находятся в различных состояниях. Например, осенью и перед распусканием листьев крахмал в таких клетках находится в изобилии, а после распускания листьев содержание его значительно уменьшается. В клетках, не сообщающихся с сосудами при помощи пор, изменение состояния запасных веществ происходит очень медленно по сравнению с изменением их в клетках, сообщающихся с сосудами порами.
Вследствие того, что клетки древесинной паренхимы тесно связаны с сосудами и притом не только анатомически, но и физиологически, сосуды нельзя рассматривать совершенно изолированно. Это — система, в которой сосуд является только местом для продвижения весьма разведенных водных растворов. Предполагают, что состав растворов, а также отчасти направление их движения регулируются деятельностью паренхимных клеток, находящихся в непосредственном контакте с сосудам или входящих с ним в контакт при посредстве звеньев клеток. В сосудах многих растений, особенно древесных, сахар содержится не только весной, но и в другие времена года и иногда в заметных количествах (например, в условиях климата Закавказья — зимой). В зависимости от ряда условий клетки древесинной паренхимы, примыкающие к сосуду, могут образовывать выросты в полость сосудов — тиллы. Тиллы с течением времени разрастаются, заполняют всю полость сосуда, оболочки их одревесневают, и они становятся полным подобием создавшей их древесинной паренхимы, отличаясь от ее клеток лишь размерами и разнообразием очертаний. В таких тиллах откладываются те же запасные пластические вещества, что и в клетках древесинной паренхимы. Таким образом, сосуд с момента образования тилл тоже становится местом отложения запасных веществ.
Иногда тиллы весьма тесно заполняют полость сосуда, сильно сжимая друг друга и приобретая исключительно толстые оболочки, пронизанные поровыми каналами.
В этих случаях клетки тилл уже не похожи на паренхиму, расположенную вне полости сосуда. В некоторых редких случаях клетки тилл имеют облик и структуру типичных каменистых клеток. Тогда, плотно забивая сосуды и наполняясь различного рода отложениями, они придают ксилеме большую твердость. Особенно велико значение тилл в процессе формирования так называемого ядра древесины. Изредка тиллы образуются и у хвойных в трахеидах и смоляных ходах.
Клетки древесинной паренхимы в направлении, параллельном длинной оси органа, редко расположены беспорядочно, особенно клетки, не соприкасающиеся непосредственно с сосудом, с его стенкой. Такие клетки образуют подобие звеньев цепи, растянутой вдоль длинной оси органа. Но вертикальные ряды, составленные из клеток древесинной паренхимы, — короткие, оканчивающиеся с обеих концов клетками с заостренными окончаниями, подобно трахеидам.
Древесинная паренхима многолетних растений по своему расположению в толще годичного слоя подразделяется на два типа: паратрахеальную, приуроченную к сосудам, и апотрахеальную, не связанную с сосудами. В пределах паратрахеального типа различают вазицентрическую, крыловидную и сомкнуто-крыловидную паренхиму. Эти термины имеют чисто описательное значение и определяют характер расположения клеток вокруг сосудов.
В пределах апотрахеального типа по расположению составляющих элементов различают диффузную, метатрахеальную и терминальную паренхимы. Диффузная паренхима на поперечных срезах обнаруживается в виде одиночных клеток, рассеянных среди клеток лучей. В продольном направлении клетки диффузной паренхимы располагаются в виде цепочек. В том случае, если несколько таких цепочек располагаются рядом, древесинная паренхима называется метатрахеальной. На поперечных срезах она имеет вид тангентальных полосок, состоящих из одного ряда клеток. У многих древесных растений с сезонным приростом древесинная паренхима развивается только к концу прироста. При этом она формируется в виде более или менее сплошного слоя вокруг кольца прироста. Такая паренхима называется терминальной.
Клетки древесинной паренхимы образуются так называемыми веретенообразными клетками камбия. Если клетка камбия непосредственно дифференцируется в паренхимную клетку, последняя имеет веретеновидную форму, напоминая элементы либриформа. Такие клетки носят название заменяющих волокон и отличаются от либриформа простыми округлыми порами и остающимся в жизнедеятельном состоянии протопластом.
Однако чаще перед образованием паренхимы клетка камбия несколько раз делится в одном направлении; возникшая цепочка клеток дифференцируется как тяжевая паренхима. Сопоставляя анатомические элементы флоэмы с анатомическими элементами ксилемы, можно убедиться, что и флоэма и ксилема составлены из элементов трех основных типов, приспособленных к выполнению соответствующих функций: запасающей, проводящей и механической. Между этими тремя основными типами существуют переходные как в структурном, так и в функциональном отношении. Так, анатомическим элементом, предназначенным преимущественно для отложения и хранения различных запасных веществ, является паренхима, лубяная во флоэме и древесинная в ксилеме. Очертания паренхимных клеток в обоих участках проводящей системы разнообразны; причем различие заключается в особенностях строения оболочек. Клетки лубяной паренхимы имеют целлюлозные оболочки, а клетки древесинной паренхимы — одревесневшие. Поры в обоих типах паренхимы простые, если протопласт клетки находится в жизнедеятельном состоянии. В клетках древесинной паренхимы с отмершим протопластом поры могут быть и окаймленными, причем степень развития и рисунок окаймленных пор у таких клеток различны.
Механическая система как во флоэме, так и в ксилеме представлена волокнами. У некоторых растений в коре встречаются паренхимные толстостенные клетки (каменистые), но в самой флоэме таких клеток не образуется; они характерны для коровых участков сердцевинных лучей.
Между лубяными волокнами флоэмы и либриформом структурная аналогия еще более полная, чем между анатомическими элементами запасающей и проводящей систем. Оболочки лубяных волокон также нередко одревесневают, уподобляясь в этом отношении либриформу. Поры у таких лубяных волокон щелевидные и косые (вариант простых пор), вполне аналогичные порам клеток либриформа.
Оболочки механических элементов флоэмы и ксилемы толстостенные. У некоторых растений как лубяные волокна, так и волокна либриформа имеют поперечные перегородки.
Вообще анатомические элементы флоэмы и ксилемы, подобно веем элементам растительных тканей, очень пластичны и представлены рядом переходных форм. Особенно много переходных
форм наблюдается в узлах стеблей — в тех местах, где от стебля отходят листья и ветки, а также в местах поранения, где происходит заживление ран.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Источник