
Если ли в сосудах нервные волокна

Оглавление темы “Вегетативная ( автономная ) нервная система.”:
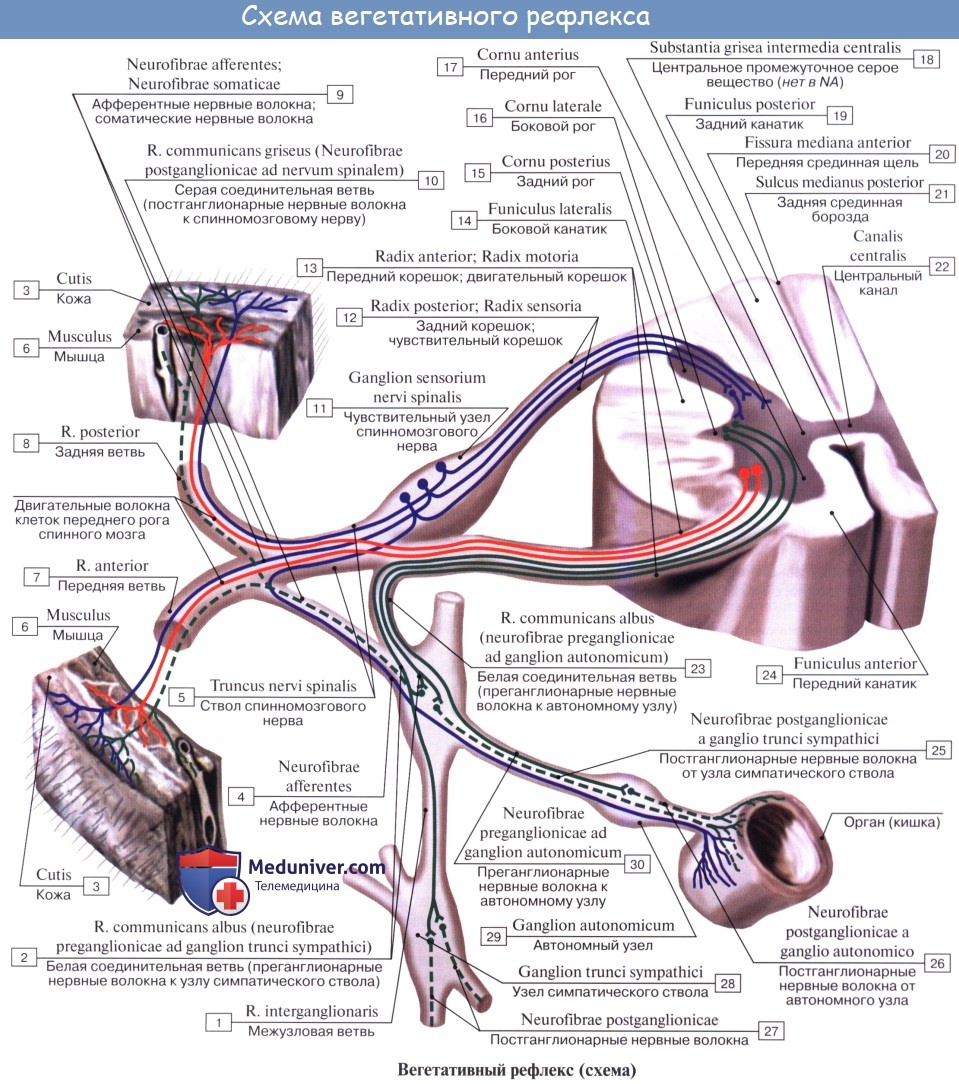
Иннервация кровеносных сосудов. Иннервация сосудов.Степень иннервации артерий, капилляров и вен неодинакова. Артерии, у которых более развиты мышечные элементы в tunica media, получают более обильную иннервацию, вены — менее обильную; v. cava inferior и v. portae занимают промежуточное положение. Более крупные сосуды, расположенные внутри полостей тела, получают иннервацию от ветвей симпатического ствола, ближайших сплетений вегетативной нервной системы и прилежащих спинномозговых нервов; периферические же сосуды стенок полостей и сосуды конечностей получают иннервацию от проходящих поблизости нервов. Нервы, подходящие к сосудам, идут сегментарно и образуют периваскулярные сплетения, от которых отходят волокна, проникающие в стенку и распределяющиеся в адвентиции (tunica externa) и между последней и tunica media. Волокна иннервируют мышечные образования стенки, имея различную форму окончаний. В настоящее время доказано наличие рецепторов во всех кровеносных и лимфатических сосудах. Первый нейрон афферентного пути сосудистой системы лежит в спинномозговых узлах или узлах вегетативных нервов (nn. splanchnici, n. vagus); далее он идет в составе кондуктора интероцептивного анализатора (см. «Интероцептивный анализатор»). Сосудодвига-тельный центр лежит в продолговатом мозге. К регуляции кровообращения имеют отношение globus pallidus, таламус, а также серый бугор. Высшие центры кровообращения, как и всех вегетативных функций, заложены в коре моторной зоны головного мозга (лобная доля), а также впереди и сзади нее. Корковый конец анализатора сосудистых функций располагается, по-видимому, во всех отделах коры. Нисходящие связи головного мозга со стволовыми и спинальными центрами осуществляются, по-видимому, пирамидными и экстрапирамидными трактами.
Замыкание рефлекторной дуги может происходить на всех уровнях центральной нервной системы, а также в узлах вегетативных сплетений (собственная вегетативная рефлекторная дуга). Эфферентный путь вызывает вазомоторный эффект — расширение или сужение сосудов. Сосудосуживающие волокна проходят в составе симпатических нервов, сосудорасширяющие волокна идут в составе всех парасимпатических нервов краниального отдела вегетативной нервной системы (III, VII, IX, X), в составе передних корешков спинномозговых нервов (признается не всеми) и парасимпатических нервов сакрального отдела (nn. splanchnici pelvini). Учебное видео вегетативной иннервации внутренних органовДругие видео уроки по данной теме находятся: Здесь – Также рекомендуем “Единство вегетативной и центральной нервной системы. Зоны Захарьина — Геда.” Редактор: Искандер Милевски. Дата последнего обновления публикации: 28.8.2020 |
Источник
Активно работающие нейроны побуждают сосуды мозга расти и ветвиться, если же нейроны молчат, сосудистая сеть делается более разреженной.
Нейроны мозга зависят от питающих их кровеносных сосудов. Известно, что во время напряжённой работы мозга усиливается кровоток в тех его областях, которые работают активнее прочих – местные клетки нужно обеспечить питательными веществами и кислородом. Как выяснили исследователи из Медицинской школы Гарварда (США), взаимосвязь между нейронами и кровеносными сосудами на самом деле ещё прочнее, чем полагали ранее – оказывается, сама структура сосудистой сети может меняться в ответ на изменения в нейрональной активности.
Участок коры мозга, принимающий сигналы от вибриссов: аксоны нейронов окрашены красным, кровеносные сосуды – зелёным. (Фото Baptiste Lacoste / Harvard Medical School).
Кровеносные сосуды в коре мозга мышей при отсутствии сенсорных сигналов (слева) и после сенсорной стимуляции (справа). (Фото Baptiste Lacoste / Harvard Medical School).
‹
›
Известно, что нейронная архитектура мозга весьма пластична – нервные клетки постоянно формируют и разрывают контакты друг с другом, а доля белого и серого вещества в разных мозговых отделах может довольно сильно меняться в зависимости от того, какие задачи приходится постоянно выполнять мозгу. Но это нейроны, насчёт же сосудистой сети считалось, что она полностью определяется генами, которые руководят её формированием в ходе индивидуального развития. В статье в Neuron её авторы признают, что гены играют тут огромную роль, особенно на первых этапах созревания мозга, однако дальнейшая настройка сосудистой системы во многом зависит от нейронной активности.
Баптисте Лакост (Baptiste Lacoste) и его коллеги экспериментировали с мышами, пометив у животных нейроны и кровеносные сосуды разными флуоресцентными белками: аксоны нейронов – красным, сосуды – зелёным, чтобы можно было наблюдать за изменениями в тех и в других. В качестве «подопытных» были выбраны нервные клетки из области коры, принимавшей сенсорные сигналы от вибриссов. Исследователей интересовало, изменится ли что-то в сосудах коры, если в неё перестанут поступать сигналы извне (для этого мышей либо модифицировали генетически – так, чтобы нервные импульсы от чувствительных нейронов не доходили до мозга, либо просто удаляли сами вибриссы). Как следствие, кровеносные сосуды в коре укорачивались и они делались менее разветвлёнными. Наоборот, когда вибриссы стимулировали, то и сосуды в вибриссной коре становились длиннее и формировали новые ответвления.
В дальнейшем нейробиологи хотят выяснить, как именно связаны нейроны и сосуды. Возможно, передачей сигналов от одних к другим занимаются астроциты, специальные вспомогательные клетки нервной системы, чья забота – следить за благополучием нейронов (и которых, кстати, в пять раз больше, чем нейронов). Также предстоит более тщательно изучить факторы, которые могут повлиять на взаимодействие нейронов и сосудистой сети. Многие заболевания сопровождаются тем, что мозг перестаёт получать в достаточном количестве кислород и питательные вещества; возможно, чтобы такого не происходило, нужно позаботиться о том, чтобы мозг не испытывал недостатка в ощущениях.
Источник
Периферические нервы отличаются высоким уровнем кровообращения, что обеспечивает интенсивный метаболизм в них. Знание общих закономерностей кровоснабжения нервов и частной микрососудистой анатомии в зоне расположения конкретных нервных стволов очень важно при выполнении пластических и реконструктивных операций на конечностях. Оно позволяет свести к минимуму нарушения кровообращения в нервах при их выделении из тканей, а значит, создать более благоприятные условия для последующей регенерации нервных волокон.
Внешняя сосудистая сеть нерва
Внешняя сосудистая сеть образована сосудами, сопровождающими нерв на большем или меньшем протяжении. Различного калибра артериальные ветви подходят к крупным нервным стволам через каждые 2—10 см. Наибольшее практическое значение имеют следующие четыре основных типа кровоснабжения нервных стволов.

Рис. 2.5.1. Схематическое изображение некоторых типов кровоснабжения крупных многопучковых нервов.
Тип 1 отличается отсутствием доминирующей артерии (рис. 2.5.1, а).
В связи с тем, что большинство периферических нервов входят в состав сосудисто-нервных пучков, отсутствие доминирующей артерии может наблюдаться лишь на сравнительно небольших по протяженности участках крупных и мелких нервных стволов. Кровоснабжение в этих зонах осуществляется через мышечно-кожные и перегородочно-кожные перфорирующие артерии. Небольшие мелкие ветви нервов могут не сопровождаться сопутствующими сосудами и снабжаться за счет связей с сосудистыми сплетениями окружающих тканей.
Тип 2 характеризуется наличием одной доминирующей артерии (рис 2.5.1, б), которая может сопровождать нерв на значительном протяжении. Это —один из частых вариантов строения внешней сосудистой сети, характерный для большеберцового, межреберных нервов, нервов кисти и стопы, срединного и лучевого нервов (на уровне плеча) и других стволов.
Тип 3 предполагает питание нерва через множественные доминирующие артерии (рис. 2.5.1, в). Этот тип кровоснабжения характерен для локтевого нерва в верхней трети предплечья, для поверхностной ветви лучевого нерва и др.
Тип 4 встречается в той зоне нерва, где участок с преобладающей артерией переходит в свободный от доминирующих сосудов участок (рис. 2.5.1, г). Этот вариант кровоснабжения характерен для лучевого и малоберцового нервов в зоне их деления на конечные ветви.
Внутриствольная сосудистая сеть
Внутриствольная сосудистая сеть образуется ветвями расположенных вблизи нервов сосудов, которые подходят к нерву и делятся на восходящую и нисходящую ветви. Их конечные разветвления, анастомозируя между собой, образуют выраженную интраневральную непрерывную сеть, ячейки которой представлены прекапиллярами и капиллярами, вытянуты между волокнами и располагаются между ними
С практической точки зрения, целесообразнс выделить два основных варианта строения интраневральной сосудистой сети. Первый из них отличается наличием внутриствольно расположенной доминирующей артерии и характерен только для крупных нервных стволов на тех участках, где они проходят вне сосудистых пучков.
Это — срединный (на предплечье) и седалищный нервы, сосуды которых располагаются субэпиневрально или внутриствольно и могут достигать 1—2 мм в диаметре. Для второго варианта характерно отсутствие интраневральной доминирующей артерии. При этом внутриствольная сеть представлена сосудами малого калибра.
Следует отметить, что в любых крупных многопучковых нервах наиболее значительные по диаметру сосуды располагаются в наружном эпииеприи, что позволяет использовать их для идентификации соответствующих нервных пучков при сшивании и пластике.
Кровообращение в нервах с позиций пластической хирургии
Как известно, успех пластических и реконструктивных операций на нервах в значительной степени зависит от того, насколько хирургу удастся сохранить кровообращение в концах нервного ствола. Описанные выше типы строения экстра- и интраневральной сосудистой сети определяют оптимальную технику подготовки концов нерва к сшиванию или пластике, которая в той или иной степени всегда связана с выделением концов нерва из окружающих тканей, т.е. с отсечением на определенном отрезке внешних источников питания.
Так, при отсутствии доминирующих артерий во внешней сосудистой сети (тип 1) внутриствольнос кровообращение в нерве обеспечивается в максимальной степени при выделении его концов из тканей вместе с прилегающей клетчаткой, когда сохраняют непрерывность сосудистой сети (рис. 2.5.2).
Рис. 2.5.2. Схематическое изображение оптимального уровня пересечения параневральных сосудов (стрелки) при отсутствии доминирующей экстраневральной артерии.
Однако клинические наблюдения показывают, что и в этом случае кровоснабжение концов нерва, выделенных на протяжении 8—10 см (или более), значительно снижается, особенно на периферическом отрезке. Эти нарушения выражены в минимальной степени, когда концы нерва выделяют вместе с доминирующей артерией. При этом протяженность участка выделения существенного значения не имеет.
Особенно просто задача выделения нерва из тканей решается при субэпиневральном расположении доминирующей артерии. При этом выделение концов нерва ограничивается прежде всего необходимостью выделения (пересечения) его ветвей.
При экстраневральном расположении доминирующей артерии (тип 2) выделение концов нерва из тканей следует по возможности осуществлять с включением сопутствующего сосудистого пучка, что сохраняет сосудистую сеть нерва практически неизменной.
При 3-м и 4-м типах строения внешней сосудистой сети нерва, когда рядом с ним на определенном участке проходит крупный сосудистый пучок (например, локтевой на предплечье или плечевой на плече), хирург может оказаться в трех различных ситуациях.
Прежде всего при сохранении целости магистральных сосудов их пересечение и выделение из тканей вместе с нервом, как правило, нецелесообразны, а часто —недопустимы. Поэтому нервный ствол выделяют так же, как и при 1-м типе его кровоснабжения.
Когда поврежден весь сосудисто-нервный пучок и когда нет необходимости в восстановлении магистральных сосудов (рис. 25.3, а), концы нерва можно выделять одним блоком с сосудами до того участка, где сосуды уходят в сторону (рис. 2 53, б). Если – же необходимо выделить нерв и более проксимально, то включать сосудистый пучок в выделяемый лоскут, как правило, нецелесообразно (рис. 2 5 3, в).

Рис. 2.5.3. Схема вариантов выделения концов нервного ствола в зависимости от расположения внешней доминирующей артерии (объяснение в тексте).
Следует отметить, что протяженность участка выделения концов нерва из тканей и техника этого этапа операции определяются не только архитектоникой сосудистой сети в зоне повреждения, но и такими факторами, как тип оперативного вмешательства (сшивание нерва, пластика, транспозиция и пр.), выраженность и распространенность рубцовых изменений окружающих тканей, наличие сопутствующих повреждений других сосудов сегмента и т. д.
А.Е. Белоусов
Опубликовал Константин Моканов
Источник
Не́рвные воло́кна — длинные отростки нейронов, покрытые глиальными оболочками. По нервным волокнам распространяются нервные импульсы, по каждому волокну изолированно, не заходя на другие[1][2].
В различных отделах нервной системы оболочки нервных волокон значительно различаются по своему строению, что лежит в основе деления всех волокон на миелиновые и безмиелиновые. Те и другие состоят из отростка нервной клетки, лежащего в центре волокна, и поэтому называемого осевым цилиндром (аксоном), и, в случае миелиновых волокон, окружающей его оболочкой.
В зависимости от интенсивности функциональной нагрузки нейроны формируют тот или иной тип волокна. Для соматического отдела нервной системы, иннервирующей скелетную мускулатуру, обладающую высокой степенью функциональной нагрузки, характерен миелиновый (мякотный) тип нервных волокон, а для вегетативного отдела, иннервирующего внутренние органы — безмиелиновый (безмякотный) тип.
Покрытые оболочкой сплетения пучков нервных волокон образуют нервы.
Классификация Эрлангера-Гассера[править | править код]
Является наиболее полной классификацией нервных волокон по скорости проведения нервного импульса.
| Тип волокна | Функция | Диаметр, мкм | Скорость проведения, м/с | Миелинизация |
|---|---|---|---|---|
| Aα | Афферентные — мышечные веретёна, сухожильные органы; эфферентные — скелетные мышцы | 10-20 | 60-120 | + |
| Aβ | Афферентные — тактильное чувство; коллатерали Aα волокон к интрафузальным мышечным волокнам | 7-15 | 40-90 | + |
| Aγ | Эфферентные — мышечные веретёна | 4-8 | 15-30 | + |
| Aδ | Афферентные — температура, быстрое проведение боли | 3-5 | 5-25 | + |
| B | Симпатические, преганглионарные; постганглионарные волокна цилиарного ганглия | 1-3 | 3-15 | прерывистая |
| C | Симпатические, постганглионарные; афферентные — медленное проведение боли | 0,3-1 | 0,5-2 | – |
Классификация по Ллойду[править | править код]
Классифицирует только афферентные нейроны.
| Тип волокна | Функция | Диаметр, мкм | Скорость проведения, м/с | Миелинизация |
|---|---|---|---|---|
| Ia | Мышечные веретёна | 18-22 | 90-120 | + |
| Ib | Сухожильные рецепторы | 15-18 | 60-90 | – |
| II | Механорецепторы кожи, вторичные мышечные веретёна | 7-15 | 40-90 | + |
| III | Рецепторы связок | 1-5 | 3-25 | прерывистая |
| IV | Болевые рецепторы, рецепторы соединительной ткани | 0,1-1 | 0,5-2 | – |
Миелинизация нервных волокон[править | править код]
При формировании безмиелинового нервного волокна осевой цилиндр (отросток нейрона) погружается в тяж из леммоцитов, цитолеммы которых прогибаются и плотно охватывают осевой цилиндр в виде муфты, края которой смыкаются над ним, образуя дупликатуру клеточной мембраны — мезаксон. Соседние леммоциты входящие в состав сплошного глиального тяжа своими цитолеммами образуют простые контакты. Безмиелиновые нервные волокна имеют слабую изоляцию, допускающую переход нервного импульса с одного волокна на другое, как в области мезаксона, так и в области межлеммоцитарных контактов.
Миелиновые нервные волокна значительно толще безмиелиновых. Принцип образования их оболочек такой же, как и безмиелиновых, то есть осевые цилиндры также прогибают цитолемму глиоцитов, образуя линейный мезаксон. Однако, быстрый рост нейронов соматического отдела нервной системы, связанный с формированием и ростом всего организма, приводит к вытягиванию мезаксонов, многократному обращению леммоцитов вокруг осевых цилиндров. В результате образуются концентрические наслоения. При этом цитоплазма с ядром леммоцитов оттесняется в область последнего витка, образующего наружный слой оболочек волокна, называемой шванновской оболочкой или неврилеммой. Внутренний слой, состоящий из витков мезаксона, называется миелиновым или миелиновой оболочкой. Следствием того, что миелинизация происходит в процессе роста как отростков нейронов, так и самих леммоцитов, является постепенное увеличение количества витков и размеров мезаксона, то есть каждый последующий виток шире предыдущего. Следовательно, последний виток, содержащий цитоплазму с ядром леммоцита является самым широким. Толщина миелина по длине волокна неоднородна, а в местах контактов соседних леммоцитов слоистая структура исчезает и контактируют лишь наружные слои, содержащие цитоплазму и ядро. Места их контактов называются узловыми перехватами (перехватами Ранвье), возникающими вследствие отсутствия здесь миелина и истончения волокна.
В ЦНС миелинизация нервного волокна происходит за счет обхвата осевых цилиндров отростками олигодендроцитов.
Как мембранная структура миелин имеет липидную основу и при обработке окисями окрашивается в тёмный цвет. Другие компоненты мембраны и промежутки не окрашиваются, поэтому периодически встречаются светлые полоски − насечки миелина (насечки Шмидта-Лантермана), которые соответствуют небольшим прослойкам цитоплазмы леммоцита.
В цитоплазме осевого цилиндра располагаются продольно ориентированные нейрофибриллы и митохондрии, которых больше в непосредственной близости к перехватам и в концевых аппаратах волокна. Цитолемма осевого цилиндра (аксона) называется аксолеммой. Она обеспечивает проведение нервного импульса, который представляет собой волну деполяризации аксолеммы. Если осевой цилиндр представлен нейритом, то в нём отсутствуют гранулы базофильного вещества.
См. также[править | править код]
- Нейрон
- Миелиновая оболочка
- Демиелинизация
- Гассер
Примечания[править | править код]
Литература[править | править код]
- Савельев А. В. Моделирование логики самоорганизации активности нервного пучка эфаптическими взаимодействиями аксонного уровня // сб.: Моделирование неравновесных систем. — Институт вычислительного моделирования СО РАН, Красноярск, 2004. — С. 142—143.
Источник