
Нервные волокна кровеносных сосудов

Оглавление темы “Вегетативная ( автономная ) нервная система.”:
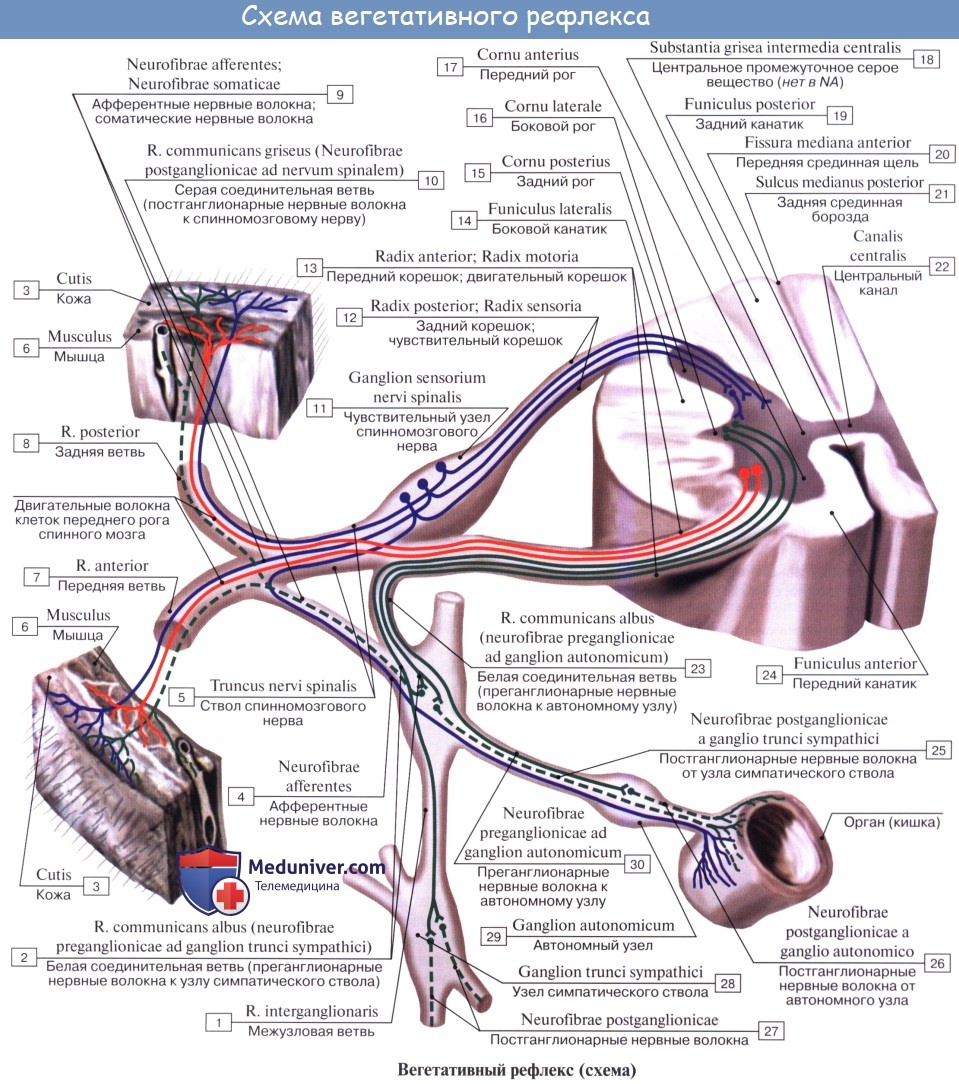
Иннервация кровеносных сосудов. Иннервация сосудов.Степень иннервации артерий, капилляров и вен неодинакова. Артерии, у которых более развиты мышечные элементы в tunica media, получают более обильную иннервацию, вены — менее обильную; v. cava inferior и v. portae занимают промежуточное положение. Более крупные сосуды, расположенные внутри полостей тела, получают иннервацию от ветвей симпатического ствола, ближайших сплетений вегетативной нервной системы и прилежащих спинномозговых нервов; периферические же сосуды стенок полостей и сосуды конечностей получают иннервацию от проходящих поблизости нервов. Нервы, подходящие к сосудам, идут сегментарно и образуют периваскулярные сплетения, от которых отходят волокна, проникающие в стенку и распределяющиеся в адвентиции (tunica externa) и между последней и tunica media. Волокна иннервируют мышечные образования стенки, имея различную форму окончаний. В настоящее время доказано наличие рецепторов во всех кровеносных и лимфатических сосудах. Первый нейрон афферентного пути сосудистой системы лежит в спинномозговых узлах или узлах вегетативных нервов (nn. splanchnici, n. vagus); далее он идет в составе кондуктора интероцептивного анализатора (см. «Интероцептивный анализатор»). Сосудодвига-тельный центр лежит в продолговатом мозге. К регуляции кровообращения имеют отношение globus pallidus, таламус, а также серый бугор. Высшие центры кровообращения, как и всех вегетативных функций, заложены в коре моторной зоны головного мозга (лобная доля), а также впереди и сзади нее. Корковый конец анализатора сосудистых функций располагается, по-видимому, во всех отделах коры. Нисходящие связи головного мозга со стволовыми и спинальными центрами осуществляются, по-видимому, пирамидными и экстрапирамидными трактами.
Замыкание рефлекторной дуги может происходить на всех уровнях центральной нервной системы, а также в узлах вегетативных сплетений (собственная вегетативная рефлекторная дуга). Эфферентный путь вызывает вазомоторный эффект — расширение или сужение сосудов. Сосудосуживающие волокна проходят в составе симпатических нервов, сосудорасширяющие волокна идут в составе всех парасимпатических нервов краниального отдела вегетативной нервной системы (III, VII, IX, X), в составе передних корешков спинномозговых нервов (признается не всеми) и парасимпатических нервов сакрального отдела (nn. splanchnici pelvini). Учебное видео вегетативной иннервации внутренних органовДругие видео уроки по данной теме находятся: Здесь – Также рекомендуем “Единство вегетативной и центральной нервной системы. Зоны Захарьина — Геда.” Редактор: Искандер Милевски. Дата последнего обновления публикации: 28.8.2020 |
Источник
Кардиогенез :: Нервы сосудов и сердца. (Курс гистологии, Заварзин, 1946)

(Заварзин А.А., Румянцев А.А. Курс гистологии. 1946г)
ГЛАВА ДЕВЯТАЯ СОСУДИСТАЯ СИСТЕМА И СОСУДИСТЫЕ ОРГАНЫ
КРОВЕНОСНЫЕ И ЛИМФАТИЧЕСКИЕ СОСУДЫ
КРОВЕНОСНЫЕ СОСУДЫ
стр.432-434
Нервы сосудов и сердца
Сосуды и сердце иннервируются волокнами вегетативной нервной системы.
Иннервация сосудов. Схема симпатической иннервации сосудов показана на рис. 372.
Можно считать доказанным, что сосудосуживающий центр цереброспинальной системы расположен в стволовой части мозга в плоскости ядер седьмого нерва. Невриты клеток этого центра идут в спинной мозг, где, в конце концов, и дают синапсы с нервами симпатических ядер. От этих последних начинаются преганглионарные волокна (рис. 372, 9), идущие в пограничный ствол, где они и заканчиваются синапсами на клетках Гольджи.
От пограничного ствола идут постганглионарные волокна (3), которые входят в стенку сосудов и образуют здесь разветвленля в форме сетей.
Постганглионарные волокна безмякотные. Импульсы, пришедшие по этим волокнам, повышают тонус мышечной ткани, и сосуд начинает сокращаться.

Чувствительные волокна представлены дендритами невронов спинальных ганглиев (2); они заканчиваются в стенках сосудов рецепторами самого различного рода, а также дают терминальные разветвления, располагающиеся в тех же местах, где находятся сплетения, образованные симпатическими волокнами. Все чувствительные волокна мякотные, но, войдя в стенку сосуда, они теряют свои оболочки и поэтому не могут быть отличимы от волокон симпатических. Войдя в спинной мозг, невриты чувствительного волокна направляются по ним в стволовую часть, где и разветвляются в области сосудосуживающего центра, а их коллатерали идут в боковые рога, где и образуют синапсы с нейронами симпатических ядер.
Предполагается, что чувствительные волокна могут быть двух родов. По одним импульсы, идущие от стенки сосуда, вызывают возбуждение сосудосуживающего центра, в результате которого тонус стенки, естественно, будет повышен. По другим импульсы, идущие в сосудосуживающий центр, вызывают его угнетение и как результат — падение тонуса сердца. Была предпринята попытка найти в сосудистой стенке рецепторы, соответствующие этим двум типам волокон, однако убедительных данных до сих пор никем ещё не представлено.
Кроме сосудосуживающих волокон, некоторые авторы допускают и наличие волокон сосудорасширяющих, относимых к парасимпатической системе, т. е. к системе блуждающего нерва. Однако сведения о них ещё более неточны, чем в отношении двух типов чувствительных волокон, а в морфологическом отношении о них вообще ничего не известно.
Нервные окончания и нервные сети в сосудистой стенке. Непосредственно под эндотелием в соединительнотканном слое всех артерий и вен располагается нервное сплетение, образованное тончайшими нервными окончаниями. Считается, что это подэндотелиальное сплетение чувствительной природы имеет самое существенное значение в передаче импульсов, возникающих в результате изменений кровяного давления. Волокна описываемого сплетения, образованного разветвлениями мякотных и безмякотных волокон, находятся в связи с другими нервными сплетениями, лежащими в адвентиции. Одно сплетение, состоящее из пучков нервных волокон, лежит более поверхностно, а отходящие от него тонкие веточки образуют второе сплетение, расположенное глубже (рис. 373), на границе со средней оболочкой.
По новейшим данным (Лаврентьев), от этого сплетения в мышечную оболочку отходят веточки, дающие окончания на отдельных мышечных клетках. Это, несомненно, концевые аппараты сосудосуживающих нервов, описанных выше.
Надо вообще помнить, что нервные сплетения сопровождают кровеносные и лимфатические сосуды на всем их протяжении.
Капилляры, как мы уже знаем, также оплетены сетью безмякотных нервов (рис. 374). По ходу сосудов, особенно более крупных, встречаются и отдельные нервные клетки, и целые их группы. Клетки эти, несомненно, симпатической природы и происходят от пограничного ствола. Изучены эти смещенные клетки ещё мало.

Несколько больше известно о чувствительных окончаниях в адвентиции сосудов: Концевые древовидные аппараты были неоднократно описаны в самом адвентиции. Кроме того, по ходу сосудов различных органов в соединительной ткани, облекающей сосуды, постоянно встречаются рецепторные аппараты от свободных древовидных окончаний до фатер-пачиниёвых телец включительно.
Из приведенного описания нетрудно заключить, что сосудистые нервы пока что изучены мало; особенно это касается способа окончания. Во всяком случае по микроскопическим картинам не только нельзя различить суживающие и расширяющие нервы, но даже не всегда удается установить, с какими волокнами мы имеем дело в каждом данном случае, — вегетативными или спинномозговыми, эффекторными или рецепторными. Дело в том, что мякотные волокна, постоянно встречающиеся в стенках сосудов, могут быть и спинальными, и вегетативными.
Иннервационные механизмы сердца. Нервная система сердца берет начало из двух источников. К сердцу подходят нервы от пограничного ствола и сердечные ветви блуждающего нерва. Волокна всех этих нервных стволов образуют в области разветвления трахеи и больших артерий два сердечных сплетения, в которых волокна настолько смешиваются, что в дальнейшем природа отдельных волокон может быть установлена только при помощи метода перерезок, а не анатомически.
В сердечных сплетениях встречаются и скопления ганглиозных клеток, образующие иногда настоящие, анатомически отличимые ганглии.
Из волокон этих сплетений уже в стенке самого сердца образуются правое и левое венечные сплетения, от которых происходят нервы, разветвляющиеся в сердце. Эти нервы образуют основное субэпикардиальное сплетение, расположенное между эпи- и миокардом и состоящее из пучков, содержащих мякотные и безмякотные волокна. Из субэпикардиального сплетения нервные волокна проникают и в миокард, и в эндокард, где также образуются сплетения. В основном сплетении сердца лежат многочисленные ганглиозные клетки, часто собранные группами в мелкие узелки. Хотя клетки эти и у человека, и у млекопитающих располагаются довольно равномерно по всему сердцу, но все-таки можно отметить, что главная их масса заложена в дорзальных стенках предсердий и у устья полых вен.
Нервные клетки имеются в области синусного узла, в перегородке предсердий, в атриовентрикулярном узле и в желудочках (в области проводящей системы). Скопления их наблюдаются и в верхней части желудочков (в продольной и венечной бороздах), а также у основания аорты и легочной артерии.
Среди клеток можно различать те же типы, которые отмечены нами при рассмотрении ганглиев автономной системы. Таким образом, всю совокупность нервных клеток сердечных сплетений и сердечной стенки можно рассматривать как диффузный ганглий автономной нервной системы.
Во время эмбрионального развития нервные клетки попадают в зачаток сердца из блуждающего нерва.
От нервных стволиков, проникающих в миокард, отходят многочисленные безмякотные веточки, густо оплетающие все перекладины сердечного мышечного синцития. Относительно способа окончания этих безмякотных нервов, являющихся, по всей вероятности, двигательными, эффекторными нервами, точных данных не имеется. Есть указания, что к отдельным мышечным перекладинам подходят безмякотные веточки, которые проникают внутрь и оканчиваются там незначительным концевым утолщением наподобие того, как оканчиваются нервы в гладких мышцах. Во всяком случае никаких образований, сколько-нибудь похожих на двигательные бляшки в скелетных мышцах, здесь не наблюдается. Чувствительные же (проприоцептивные) окончания эпилеммального типа найдены в новейшее время (Лаврентьев) в сердечной мышце в большом количестве.
Как в толстых прослойках соединительной ткани между пучками миокарда, так равно и в эпикарде, и в эндокарде встречаются разнообразные рецепторные концевые аппараты. Особенно много их в эндокарде. Здесь имеются и свободные окончания – как древовидные, так и клубочковые, и инкапсулированные аппараты в виде клубочков и телец Гольджи-Маццони.
Источник
Структурно-функциональная единица нервной системы – нервная клетка с ее отростками.
Трофическим центром клетки является тело (перикарион); воспринимающие (центрипетальные)
отростки носят название дендритов. Отросток, по которому нервный импульс идет
центрифугально, от тела клетки к рабочему органу, обозначается как аксон (нейрит).
Нервное волокно состоит из аксона (нейрита, осевого цилиндра) и окружающих его
шванновских клеток (леммоцитов), образующих неврилемму. В мякотных (миелинизированных)
нервных волокнах кнаружи от миелинового слоя располагается неврилемма или шванновская
оболочка. На относительно правильных промежутках миелиновая обкладка прерывается
и нервное волокно разделяется на сегменты. Каждый сегмент образован одним леммоцитом.
Между сегментами имеются промежутки, в которых отсутствует миелиновая оболочка
(перехваты Ранвье); именно в этих местах активно происходят обменные процессы,
способствующие проведению нервного импульса по аксону.
Нервный ствол и его ветви составлены из аксонов, берущих начало от тел клеток
нескольких типов, связанных с различными эффекторными и сенсорными органами
и функциями. Двигательные волокна от клеток передних рогов спинного мозга и
гомологичных ядер мозгового ствола составляют основную массу передних спинальных(и
черепных двигательных) корешков, но в них представлены также симпатические и
парасимпатические волокна. Задние корешки спинного мозга и чувствительные –
мозгового ствола, – содержат сенсорные волокна, тела клеток которых заключены
в ганглиях задних корешков (межпозвонковых узлах) и гомологичных ганглиях головного
мозга. После соединения спинальных корешков формируются функционально смешанные
нервные фуникулы (канатики Сикара), а затем, – на шейном, грудном, поясничном
и крестцовом уровнях – сплетения. Из этих сплетений образуются крупные нервные
стволы, несущие моторные и сенсорные волокна. Таким образом, не касаясь пока
черепных нервов, можно резюмировать, что к периферической спинальной («анимальной»)
нервной системе, кроме тел клеток серого вещества спинного мозга, относятся
передние и задние корешки, корешковый нерв Нажотта (от линии твердой мозговой
оболочки до спинального ганглия), спинальный ганглий (под которым расположен
передний корешок), далее после ганглия – спинальный канатик Сикара (фуникул),
который делится на задние ветви, иннервирующие затылочные и спинные мышцы и
кожу задней поверхности шеи и спины, и передние ветви, иннервирующие мышцы и
кожу вентральных отделов туловища и конечностей. С точки зрения топической классификации
заболеваний периферической нервной системы эти сведения хорошо поясняет старая
схема, предложенная Сикаром. Она же отражает и рутинные представления того времени
о почти исключительно инфекционно-воспалительном происхождения заболеваний периферической
нервной системы.
Источником симпатической иннервации на шейно-грудном уровне являются тела
нейронов в боковых рогах серого вещества спинного мозга, от которых идут прегангглионарные
миелинизированные волокна, покидающие передние корешки и контактирующие затем
с паравертебральными симпатическими ганглиями (симпатический ствол) или входящие
в состав черепных нервов. Подобно этому, преганглионарные парасимпатические
волокна идут из передних спинальных корешков в область таза, а на краниальном
уровне входят в состав III, IX и X пар черепных нервов. Парасимпатические ганглии
расположены в связанных с ними эффекторных органах или вблизи от них.
Многие крупные черепные и спинальные нервы идут в тесном продольном соприкосновени
с артериями и венами, образуя нервно-сосудистые пучки, и этот факт приходится
учитывать, имея в виду возможность вторичного поражения нервов при патологии
сосудов. На конечностях, по направлению к периферии, нервы находятся в более
тесном контакте с венами, нежели с артериями и здесь также возможно вторичное
страдание нервов (например, при варикозе, флеботромбозе), причем именно поверхностно
расположенных чувствительных ветвей нервов.
При осмотре невооруженным глазом нерв выглядит как белая шнуроподобная структура
с довольно гладкой поверхностью, покрытой плотно прилегающей, но не спаянной
с нервом, жировой тканью. В наиболее мощных нервах, таких как седалищный, через
нее просвечивают крупные нервные пучки – фасцикулы. На поперечном гистологическом
срезе наружная поверхность нерва окружена соединительнотканным футляром – периневрием,
состоящим из концентрических слоев жировых клеток, разделенных слоями коллагена.
Наконец, эндоневрий также представляет собой футляр, содержащий нервные волокна,
шванновские клетки (леммоциты), кровеносные сосуды вместе с пучками тонких эндоневральных
коллагеновых волокон, ориентированных вдоль нервных пучков. В эндоневрии содержится
также небольшое количеств офибробластов.. Эндоневральный коллаген плотно прилегает
к поверхности каждого нервного пучка.
Несомненно, что три указанных выше футляра выполняют роль механической защиты
нерва от повреждений, однако эндоневральная соединительная ткань выполняет и
роль своеобразной полупроницаемой перегородки, через которую из кровеносных
сосудов к шванновским клеткам и нервным волокнам диффундируют питательные вещества.
Окружающие нервные волокна пространства, как и гематоэнцефалический барьер,
также является барьером. Барьер «кровь-нерв» не пропускает чужеродные белковосвязанные
соединения. Продольное расположение эндоневрального коллагена имеет существенное
значение в качестве фактора, препятствующего тракционной травматизации нерва.
В то же время коллагеновый каркас допускает определенную свободу смещения нервного
волокна при сгибательных движениях конечностей и ориентирует направление роста
нервных волокон при регенерации нерва.
Структура нервных волокон неоднородна. Большинство нервов содержит миелинизированные
и немиелинизированные или слабо миелинизированные волокна с неодинаковым соотношением
их между собой. Клеточный состав эндоневральных пространств отражает уровень
миелинизации. В норме 90% обнаруживаемых в этом пространстве клеточных ядер
относится к клеткам Шванна (леммоцитам), а остальные принадлежат фибробластам
и капилярному эндотелию. При 80% шванновских клеток окружают немиелинизированных
аксоны; рядом с миелинизированными волокнами их количество уменьшено в 4 раза.
Тотальный диаметр нервного волокна, т. е. аксон-цилиндра (нейрита) и миелинового
футляра, вместе взятых, имеет не только морфологический интерес. Миелинизированные
волокна большого диаметра проводят импульсы в значительно более быстром темпе,
чем слабо миелинизированные или немиелинизированные. Наличие такой корреляции
послужило основой для создания ряда морфолого-физиологических классификаций.
Так, Warwick R,. Williams P. (1973) выделяют три класса волокон: А, В и С. А-волокна
– соматические афферентные и афферентные миелинизированные нервные волокна,
В-волокна – миелинизированные преганглионарные вегетативные волокна, С-волокна
– немиелинизированные вегетативные и сенсорные волокна. А. Paintal (1973) модифицировал
эту кассификацию с учетом функциональных особенностей волокон, их размеров и
скорости проведения импульсов.
Класс А (миелинизированные волокна), афферентные, сенсорные.
Группа I. Волокна размером более 20 мкм в диаметре, со скоростью проведения
импульса до 100 м/сек. Волокна этой группы несут импульсы от рецепторов мышц
(мышечных веретен, интрафузальных мышечных волокон) и рецепторов сухожилий.
Группа II. Волокна размером от 5 до 15 мкм в диаметре, со скоростью проведения
импульсов от 20 до 90 м/сек. Эти волокна несут импульсы от механорецепторов
и вторичных окончаний на мышечных веретенах интрафузальных мышечных волокон.
Группа III. Волокна размером от 1 до 7 мкм в диаметре, со скоростью проведения
импульса от 12 до 30 м/сек. Функцией этих волокон является болевая рецепция,
а также иннервация волосяных рецепторов и сосудов.
Класс А (миелинизированные волокна), эфферентные, двигательные.
Альфа-волокна. Более 17 мкм в диаметре, скорость проведения импульса от 50
до 100 м/сек. Они иннервируют экстрафузальные поперечнополосатые мышечные волокна,
преимущественно стимулируя быстрые сокращения мышц (мышечные волокна 2-го типа)
и крайне незначительно – медленные сокращения (мышц 1-го типа).
Бета-волокна. В отличие от альфа-волокон иннервируют мышечные волокна 1-го
типа (медленные и тонические сокращения мышц) и частично интрафузальные волокна
мышечного веретена.
Гамма-волокна. Размером 2-10 мкм в диаметре, скорость проведения импульса
10-45 см/сек, иннервирует только интрафузальные волокна, т. е. мышечное веретено,
тем самым участвуя в спинальной саморегуляции мышечного тонуса и движений (кольцевая
связь гамма-петли).
Класс В – миелинизированные преганглионарные вегетативные.
Это небольшие нервные волокна, около 3 мкм в диаметре, со скоростью проведения
импульса от 3 до 15 м/сек.
Класс С – немиелинизированные волокна, размерами от 0,2 до 1,5 мкм в диаметре,
со скоростью проведения импульса от 0,3 до 1,6 м/сек. Этот класс волокон состоит
из постганглионарных вегетативных и эфферентных волокон, преимущественно воспринимающих
(проводящих) болевые импульсы
Очевидно, что эта классификация интересует и клиницистов, помогая понять некоторые
особенности эфферентной и сенсорной функций нервного волокна, в том числе закономерности
проведения нервных импульсов, как в норме, так и при различных патологических
процессах.
Электрофизиологические исследования показывают, что в состоянии покоя существует
разница в электрическом потенциале на внутренней и внешней сторонах неврональной
и аксональной клеточной мембраны. Внутренняя часть клетки имеет отрицательный
разряд 70-100 мВ по отношению к интерстициальной жидкости снаружи клетки. Этот
потенциал поддерживается различием в концентрации ионов. Калий (и белки) преобладают
внутри клетки, в то время как ионы натрия и хлориды имеют более высокую концентрацию
вне клетки. Натрий постоянно диффундирует в клетку, а калий имеет тенденцию
выходить из нее. Дифференциал концентрации натрий-калий поддерживается путем
энергозависимого насосного механизма в покоящейся клетке, причем это равновесие
существует при слегка сниженной концентрации положительно заряженных ионов внутри
клетки, чем снаружи от нее. Это приводит к отрицательному внутриклеточному заряду.
Ионы кальция также вносят вклад в поддержание равновесия в клеточной мембране,
и когда их концентрация снижается, возбудимость нерва нарастает.
Под влиянием естественной или вызванной внешними факторами стимуляции аксона
происходит нарушение селективной проницаемости клеточной мембраны, что способствует
проникновению ионов натрия в клетку и редукции потенциала покоя. Если мембранный
потенциал снижается (деполяризуется) до критического уровня (30-50 мВ), то возникает
потенциал действия и импульс начинает распространяться вдоль клеточной мембраны
как волна деполяризации. Важно отметить, то в немиелинизированных волокнах скорость
распространения импульса прямо пропорциональна диаметру аксона, и возбуждение
длительно прямолинейно захватывает соседствующие мембраны.
Проведение же импульса в миелинизированных волокнах совершается «сальтаторно»,
т. е. как бы скачкообразно: импульс или волна деполяризации мембраны скользит
от одного перехвата Ранвье до другого и так далее. Миелин действует как изолятор
и предупреждает возбуждение мембраны клетки аксона, за исключением промежутков
на уровне перехватов (узлов) Ранвье. Нарастание проницаемости возбужденной мембраны
этого узла для ионов натрия вызывает ионные потоки, которые и являются источником
возбуждения в области следующего перехвата Ранвье. Таким образом, в миелинизированых
волокнах скорость проведения импульса зависит не только от диаметра аксона и
толщины миелинового футляра, но и от дистанции между узлами Ранвье, от «интернодальной»
длины.
Большинство нервов имеет смешанный состав нервных волокон по их диаметру,
степени миелинизации (миелинизированные и немиелинизированные волокна), включение
вегетативных волокон, дистанциям между перехватами Ранвье, и поэтому каждый
нерв имеет свой, смешанный (сложный) потенциал действия и суммированную скорость
проведения импульса. Например, у здоровых лиц скорость проведения по нервному
стволу, измеренная при накожном наложении электродов, варьирует от 58 до 72
м/сек для лучевого нерва и от 47 до 51 м/сек для малоберцового нерва (M. Smorto,
J. Basmajian, 1972).
Информация, передаваемая по нерву, распространяется не только стереотипными
электрическими сигналами, но и с помощью химических передатчиков нервного возбуждения
– медиаторов или трансмиттеров, освобождаемых в местах соединения клеток – синапсах.
Синапсы – специализированные контакты, через которые осуществляется поляризованная,
опосредованная химически, передача из нейрона возбуждающих или тормозящих влияний
на другой клеточный элемент. В дистальной, терминальной части нервное волокно
лишено миелина, образуя терминальную арборизацию (телодендрон) и пресинаптический
терминальный элемент. Этот элемент морфологически характеризуется расширением
окончания аксона, что напоминает булаву и нередко именуется как пресинаптический
мешок, терминальная бляшка, бутон, синаптический узелок. Под микроскопом в этой
булаве можно увидеть различных размеров (около 500 А) гранулярные пузырьки или
синаптические везикулы, содержащие медиаторы (например, ацетилхолин, катехоламины,
пептидные гормоны и др.).
Подмечено, что присутствие круглых пузырьков отвечает возбуждению, а плоских
– торможению синапса. Под терминальной бляшкой лежит синаптическая щель размерами
0,2-0,5 мкм в поперечнике, в которую из везикул поступают кванты медиатора.
Затем следует субсинаптическая (постсинаптическая) мембрана, воздействуя на
которую химический передатчик вызывает изменения электрического потенциала в
подлежащих клеточных элементах.
Можно назвать по крайней мере две главные функции нейрона. Одна из них – поддержание
собственной функциональной и морфологической целостности и тех клеток организма,
которые данным нейроном иннервируется. Эту функциональную роль нередко обозначают
как трофическую. Вторая функция представлена сочетанием механизмов, дающих начало
возбуждению, его распространению и целенаправленной деятельности по интеграции
с другими функционально-морфологическими системами. Метаболическая зависимость
аксона от тела клетки (перикариона) была продемонстрирована еще в 1850 году,
Валлером, когда после пересечения нерва наступала дегенерация в его дистальной
части («валлеровское перерождение»). Уже само по себе это указывает на то, что
в теле нейрона имеется источник клеточных компонентов, вырабатываемых нейронным
перикарионом и направляемых вдоль аксона к его дистальному концу. Сказанное
относится не только к выработке и продвижению по нейрону к симпатической щели
ацетилхолина и других медиаторов. Электронномикроскопическая и радиоизотопная
техника позволила уточнить и новые особенности центрифугального аксоплазматического
транспорта. Оказалось, что клеточные органеллы, такие как митохондрии, лизосомы
и везикулы передвигаются по аксону с медленной скоростью 1-3 мм в день, в то
время как отдельные белки – 100 мм в день. Гранулы, аккумулирующие катехоламины,
в симпатических волокнах двигаются со скоростью от 48 до 240 мм в день, а нейросекреторные
гранулы по гипоталамо-гипофизарному тракту – 2800 мм в день. Имеются доказательства
и ретроградного аксоплазматического транспорта. Такой механизм обнаружен по
отношению к вирусам герпеса простого, возбудителям ботулизма и столбняка.
Кровеносные сосуды нервов являются ветвями близрасположенных сосудов. Подходящие
к нерву артерии разделяются на восходящую и нисходящую ветви, которые распространяются
по нерву. Артерии нервов анастомозируют между собой, образуя непрерывную сеть
по ходу всего нерва. Наиболее крупные сосуды расположены в наружном эпиневрии.
От них отходят ветви в глубину нерва и проходят в нем между пучками в рыхлых
прослойках внутреннего эпиневрия. От этих сосудов ветви проходят к отдельным
пучкам нерва, располагаясь в толще периневральных влагалищ. Тонкие ветви этих
периневральных сосудов расположены внутри пучков нервных волокон в прослойках
эндоневрия (эндоневральные сосуды). Артериолы и прекапилляры вытянуты по ходу
нервных волокон, располагаясь между ними.
По ходу седалищного и срединного нерва обычно расположены заметные и достаточно
длинные артерии (артерия седалищного нерва, артерия срединного нерва). Эти собственные
артерии нервов анастомозируют с ветвями близрасположенных сосудов.
Количество источников кровоснабжения каждого нерва индивидуально различно.
Большей или меньшей величины артериальные веточки подходят к крупным нервам
через каждые 2-10 см. В связи с этим выделение нерва из окружающей его околонервной
клетчатки в какой-то мере сопряжено с повреждением подходящих к нерву сосудов.
Микроваскулярное кровоснабжение нерва, исследованное прижизненным микроскопическим
методом показало, что обнаруживаются эндоневральные анастомозы между сосудами
в различных слоях нерва. При этом преобладает наиболее развитая сеть внутри
нерва. Изучение эндоневрального кровотока имеет большое значение как показатель
степени повреждения нерва, при этом кровоток претерпевает немедленные изменения
даже при слабой компрессии в эксперименте на животных и на людях, производимой
на поверхности нерва или же если компремируются экстраневральные сосуды. При
такой экспериментальной компрессии только часть глубокорасположенных в нерве
сосудов сохраняют нормальный кровоток (Lundborg G,. 1988).
Вены нервов формируются в эндоневрии, периневрии и эпиневрии. Наиболее крупными
венами являются эпиневральные. Вены нервов впадают в близрасположенные вены.
Следует отметить, что при затруднениях венозного оттока вены нервов могут расширяться,
образуя варикозные узлы.
Лимфатические сосуды нерва. В эндоневрии и в периневральных футлярах имеются
лимфатические щели. Они находятся в связи с лимфатическими сосудами в эпиневрии.
Отток лимфы от нерва происходит по лимфатическим сосудам, тянущимся в эпиневрии
вдоль нервного ствола. Лимфатические сосуды нерва впадают в близрасположеные
крупные лимфатические протоки, которые идут к регионарным лимфатическим узлам.
Межтканевые эндоневральные щели, пространства периневральных влагалищ являются
путями перемещения внутритканевой жидкости.
Смотрите также:
У нас также читают:
Источник