
Передвижение воды по сосудам растений

- Главная
- Природа
- Проводящая система растений
Елена Голец30 Август 2017 6653
Проводящая система растений включает в себя такие понятия, как поступление, движение воды в растениях и испарение ее. Вода необходима растениям.
Проводящая система растений
Передвигается вода в растениях по клеткам коровой паренхимы до центрального цилиндра корня, затем по проводящей системе до листовой паренхимы и, наконец, по клеткам листовой паренхимы. На первом участке пути вода передвигается осмотически благодаря повышению сосущей силы клеток корня.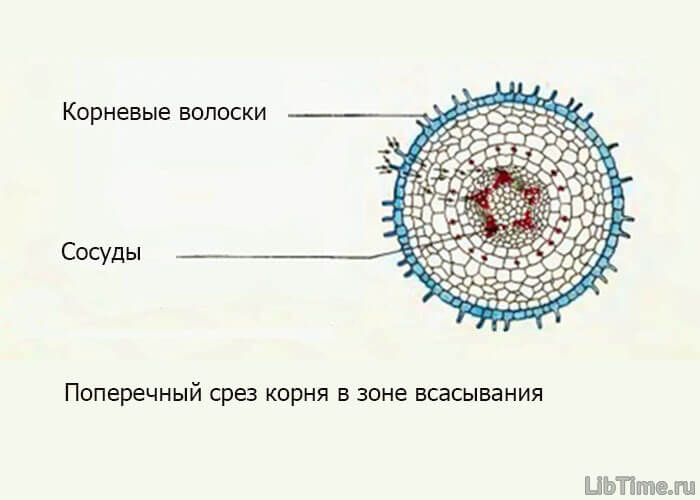 Движение воды от корневого волоска в сторону центральных сосудов.
Движение воды от корневого волоска в сторону центральных сосудов.
Этот отрезок пути очень небольшой (доли миллиметра), но передвижение воды по этому участку очень затруднено, так как воде приходится преодолевать сопротивление слоев живой протоплазмы. Это сопротивление примерно равно 1 атм на 1 мм пути, поэтому передвижение воды по живым клеткам на более значительные расстояния не обеспечивало бы потребности растения в воде.
Действительно, растения, у которых не развита проводящая система, например мхи, (подробнее: Как образуется болото) имеют незначительные размеры и приспособлены к жизни только во влажных условиях. У наземных растений в процессе эволюции образовалась проводящая ткань, которая устанавливает сообщение между всасывающими воду корнями и испаряющими воду листьями.
Проводящая воду ткань
Проводящая воду ткань состоит из сосудов, или трахей, и трахеидов; она начинается в центральном цилиндре корня, проходит через весь корень и стебель и заканчивается в виде тончайших разветвлений — жилок, пронизывающих всю листовую паренхиму.
Сосуды представляют собой мертвые трубки, образовавшиеся из живых клеток. В сосудах сохраняются поперечные перегородки на разном расстоянии (от нескольких миллиметров до метра в зависимости от вида растения) одна от другой.
Исчезновение перегородок даже на небольшом расстоянии в тысячи раз ускоряет передвижение воды. Трахеиды это длинные мертвые клетки с заостренными концами. При образовании сосудов и трахеид происходит утолщение и одревеснение их оболочек, вследствие чего они не сдавливаются под давлением окружающих их живых паренхимных клеток.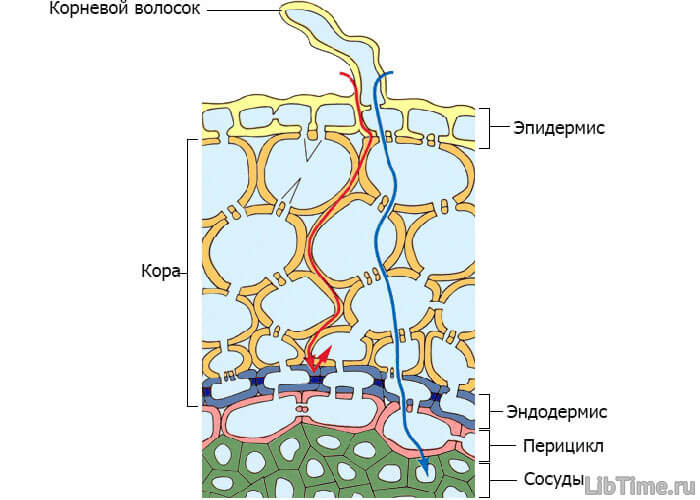 Движение воды в древесных растениях.
Движение воды в древесных растениях.
Одревеснение, однако, никогда не бывает сплошным: на стенке сосудов остаются тонкие места — поры, по которым вода может перемещаться не только вверх по сосудам, но и в радиальном направлении.
Подъем воды по сосудам
Подъем воды по сосудам можно доказать следующим опытом. Если у срезанной и поставленной в воду ветки снять кольцо коры выше уровня воды, листья ее не завянут, так как сосуды расположены в древесине.
Движение воды по сосудам чаще всего направлено снизу вверх и называется поэтому восходящим током.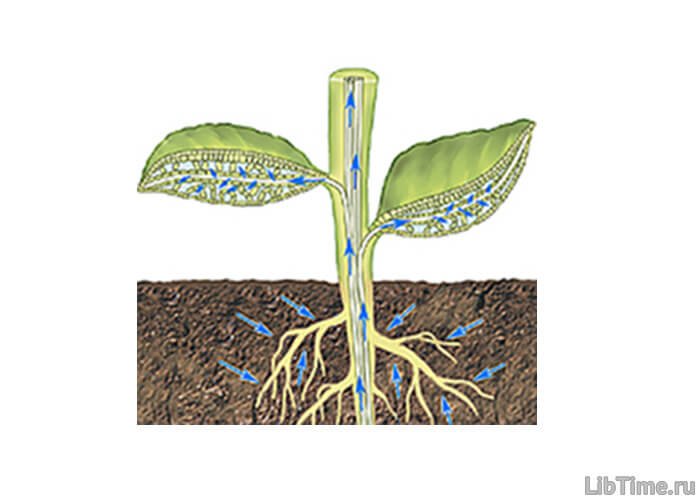 Восходящий ток.
Восходящий ток.
Последний отрезок пути водного тока по листовой паренхиме идет по живым
клеткам. Вода передвигается осмотическим путем по клеткам мезофилла листа до последних клеток, граничащих с межклеточниками. Этот отрезок пути, так же как и первый, очень короткий.
Если срезанную ветку растения герметически закрепить в стеклянной трубке, заполненной водой, и нижний конец ее опустить в сосуд со ртутью, то при испарении веткой воды ртуть в трубке будет подниматься.
Из этого опыта ясно, что передвижение воды по растению обусловлено главным образом транспирацией, (подробнее: Транспирация у растений), а не только корневым давлением.
При испарении воды с поверхности листьев в клетках возникает сосущая сила. Величина ее тем больше, чем меньше воды остается в клетках листа. Эта возникающая сосущая сила поддерживает постоянное передвижение воды в растении.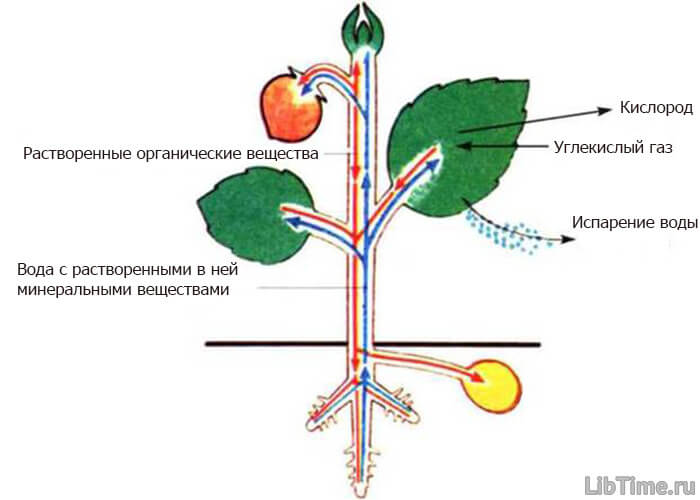 Транспорт веществ в растениях.
Транспорт веществ в растениях.
Силы, приводящие воду в движение
Таким образом, силы, приводящие воду в движение, находятся по концам проводящей системы: нагнетающий воду корень, работа которого получила название нижнего концевого двигателя, и сила присасывания воды листьями — верхний концевой двигатель.
Оба двигателя действуют в одном направлении и могут заменять и дополнять друг друга. Во время сильной инсоляции летом и при, засухе водоснабжение растения идет за счет присасывающего действия транспирации.
Корневое давление
Когда же почва богата водой, а воздух водяными парами, подъем воды обеспечивается силой корневого давления, (подробнее: Корни обеспечивают растение водой). Следовательно, в зависимости от условий внешней среды главная роль принадлежит то одному, то другому концевому двигателю.
Водные нити не рвутся под влиянием своей тяжести, несмотря на то, что при сильной транспирации они находятся в состоянии натяжения. Это объясняется силой сцепления молекул воды, достигающей 300—350 атм, а так как в сосудах нет воздуха, то целостность водного тока не прерывается.
Скорость водного тока
Скорость водного тока зависит от строения проводящих воду элементов. Вода быстрее передвигается по сосудам, причем скорость движения ее зависит от диаметра сосудов: чем он меньше, тем медленнее будет передвигаться вода.
Движение воды в растениях происходит благодаря работе двух концевых двигателей, верхнего и нижнего, и сил сцепления, обеспечивающих целостность водных нитей.
Рейтинг: 3/5 – 1
голосов
Источник
Хотя представители флоры могут абсорбировать воду всеми частями тела, основным её источником является почва. Но как она попадает от корня к вершине дерева, высотой с десятиэтажный дом? У растений нет мышц, они не могут качать жидкость по системе кровообращения. Тем не менее, вода движется по растениям вверх через клеточные стенки между протопластом клеток, через плазмодесмы (цитоплазматические мостики), через плазматические мембраны, а также через соединённые между собой проводящие элементы, распространённые по всему растению.
Сначала вода входит в корни, потом перемещается по ксилеме – внутренней проводящей ткани растения. Она поднимается, преодолевая силу тяжести Земли, благодаря двум двигателям: нижнему – корневому давлению и верхнему – испарению воды, и покидает растение в парообразном состоянии главным образом через устьица листьев (транспирация).
Большую часть пути наверх молекулы воды проходят по ксилеме. Например, по проводящим путям секвойи раствор может подниматься на высоту до 100 м. Главная роль в передвижении воды вверх принадлежит притягивающей силе, возникающей в результате испарения. При этом молекулы воды сцепляются водородными связями и движутся по стенкам трахеид и сосудов ксилемы (адгезия).
 Части растения, участвующие в подъёме воды
Части растения, участвующие в подъёме воды
Осмос усиливается аквапоринами
Вода в корневую систему поступает в зоне всасывания, через корневые волоски. Механизмы проникновения её в клетки подчиняются общим законом транспорта воды через плазмалемму. Если одиночную клетку поместить в воду, то концентрация ионов внутри ячейки будет больше, чем снаружи неё. И вода станет двигаться в клетку путём осмоса.
Однако скорость осмоса через мембрану ограничена. Долгое время учёные не могли понять, как вода может двигаться быстрее, чем предусматривает скорость осмоса. Теперь мы знаем, что осмос усиливается мембранными водными каналами, которые формируют интегральные мембранные протеины, называемые аквапоринами. Эти каналы есть в клетках животных и растений. Они проходят через мембраны вакуолей и клеточные мембраны и обеспечивают объёмный поток жидкостей.
Мембранные водные каналы ускоряют движение воды по ксилеме, обеспечивают постоянство водного баланса клетки, но они не способны изменить направление потока.

Вода движется по растениям вверх благодаря разности собственного потенциала
Направленное движение воды через плазмалемму обеспечивает разность потенциалов воды в корне и на поверхности устьиц. Потенциал воды – это вид свободной энергии. Именно градиент водного потенциала является решающей силой в определении направления движения жидкости. Жидкость движется от места большей концентрации к тому месту, где она меньше. Потенциал воды измеряется в единицах, называемых мегапаскалями (МПа).
 Корневое питание растения
Корневое питание растения
Как образуется водный потенциал?
Клеточные стенки сдерживают внутреннее давление клетки, когда вода наполняет её. Если клетку поместить в гипертонический раствор (с очень высокой концентрацией сахарозы), вода будет выходить из клетки, а клеточное давление упадёт. Клеточная мембрана отходит от клеточной стенки по мере уменьшения объёма клетки. Когда давление падает до 0, большинство растений вянут.
 Тургор и плазмолиз в клетке растений
Тургор и плазмолиз в клетке растений
Изменение размера тургора можно предсказать путём вычисления потенциала воды в клетке и окружающем растворе. Водный потенциал имеет две составляющие:
- физические силы, такие как гравитация и давление на клеточную стенку;
- концентрация растворённого вещества внутри клетки и снаружи.
Вода всегда движется в направлении более низкого потенциала воды. Например, водопад движется вниз, потому что гравитация является для него основным фактором, а потенциал в нижней части водопада ниже, чем в верхней.
На уровне клетки вклад гравитации в потенциал воды настолько мал, что обычно не входит в расчёты, если не рассматривать очень высокое дерево. Тургорное давление (давление на клеточные стенки) называется потенциалом давления. Как только увеличивается тургор, увеличивается и потенциал давления.
Концентрация растворённых веществ также определяет потенциал воды и называется потенциалом растворённого вещества. В чистой воде он нулевой. Когда в ней растворяют вещества, молекулы воды образуют с ними водородные связи. Становится меньше свободных молекул воды, что уменьшает водный потенциал. Раствор с большей концентрацией веществ имеет меньший потенциал.
Общий водный потенциал растительной клетки – это сумма потенциала её давления и потенциала растворённого вещества. Когда общий потенциал энергии воды внутри и снаружи клетки одинаковый, то вода не движется.

Источник
Путь воды в растении распадается на три различные по физиологии, строению и протяженности части: по живым клеткам корня; по мертвым элементам ксилемы корня, стебля, черешка и жилок; по живым клеткам листа до испаряющей поверхности.
Большая часть этого пути приходится на долю водопроводящей системы, состоящей из мертвых полых сосудов у покрытосеменных и трахеид у голосеменных растений. У травянистых растений эта часть водного пути достигает десятков сантиметров, а у древесных – многих метров.
Передвижение воды по сосудам ксилемы, длина которых может достигать нескольких десятков сантиметров, происходит довольно легко. Путь через трахеиды более трудный: от одной трахеиды к другой вода проходит через окаймленные поры; очевидно, что движение воды через них испытывает большее сопротивление, чем через сосуды. Измерения показывают, что у лиственных древесных растений проводимость древесины в 3 – 6 раз выше, чем у хвойных. В целом этот путь вода преодолевает куда легче, чем первый и третий через несколько миллиметров или даже долей миллиметров живых клеток – от корневых волосков до сосудов центрального цилиндра и от сосудов, расположенных в жилках листа, до испаряющих клеток мезофилла.
По сосудам и трахеидам вода передвигается, как по полым трубкам, подчиняясь общим гидродинамическим законам, по живым же клеткам корня и листа – осмотическим путем, с помощью разности сосущих сил соседних клеток в правильно возрастающей последовательности. Значительное сопротивление току воды при переходе ее от одной живой клетки к другой делает этот способ совершенно непригодным для передвижения воды на большое расстояние. Поэтому возникновение трахеид у папоротниковидных растений явилось важным этапом в эволюции растительного мира. Еще более совершенной стала водопроводящая система с появлением настоящих сосудов у покрытосеменных растений.
Через растение перекачивается огромное количество воды. С 1 га посевов пшеницы за лето испаряется около 2 тыс. т, клевера – 7,5, капусты – 8 тыс., Т.е. если собрать всю воду, расходуемую 1 га клевера или капусты, то получится водный бассейн площадью 1 га и глубиной 75- 80 см. Еловые молодняки южной тайги за год тратят 4,5 тыс. т воды с 1 га лесной площади, сосновые – 5,0, ольховые – до 11 тыс. т.
Видно, что леса расходуют не меньше, а даже больше, чем некоторые сельскохозяйственные культуры. эти колоссальные расходы восполняются за счет деятельности корневых систем, с достаточной быстротой всасывающих воду из почвы. На этом основана осушающая роль леса в условиях заболачивающихся лесных почв. Поддержанию водного баланса служат и хорошо развитая проводящая воду система, без задержки подающая воду к листьям, а также наличие покровных тканей, защищающих растение от излишней потери воды.
Каковы те силы, которые осуществляют непрерывный ток воды от корней через стебель и листья? У травянистых низкорослых растений механизм перетекания ксилемного сока понять легко. Корневое давление нагнетает воду в сосуды центрального цилиндра корня, а сосущие силы, возникающие в листьях благодаря процессу транспирации, притягивают эту воду. Тем самым создается постоянный ток воды по всему растению.
Процесс подъема воды от корней до листьев носит название восходящего тока, в отличие от нисходящего тока органических веществ от листьев к корням. Корневое давление, создающееся благодаря метаболизму корневых окончаний, получило название нижнего концевого двигателя водного тока. Притягивающие же воду сосущие силы листьев называют верхним концевым двигателем водного тока.
Труднее объяснить непрерывность водного столба у гигантов растительного мира – эвкалиптов, секвой и некоторых других древесных растений, высота которых достигает 140 м. Наши обычные деревья также имеют довольно большие размеры: береза – до 25 м, дуб – 40 м, сосна и ель – до 50 м. К этому следует добавить и значительную протяженность водопроводящей системы корней.
Водный ток испытывает и преодолевает силу земного притяжения, силу тяжести. В силу этого, например, обычные поршневые насосы не могут поднять воду с глубины более 10 м, ибо этот 10-метровый столб воды соответствует давлению в 1 атм. Кроме того, движение воды по ксилеме испытывает довольно значительное сопротивление, особенно у представителей голосеменных древесных растений.
Объяснение того, что водный столб протяженностью многие десятки метров не разрывается, находим в теории сцепления (когезии) и смачивания стенок сосудов и трахеид водой (адгезии). Действительно, между молекулами передвигающейся воды существуют значительные силы сцепления, заставляющие эти молекулы следовать друг за другом. Этому способствует и то, что водопроводящие элементы представляют как бы единое целое с водным потоком, так как стенки их полностью смочены, насыщены водой. В них нет воздуха. В таком состоянии они оказывают минимальное сопротивление движущемуся потоку. Кроме того, само строение сосудов не способствует передвижению пузырьков воздуха из одного сосуда в другой.
Все это весьма сильно отличает условия, создающиеся в дереве, от условий в поршневых насосах. В последних между стенками цилиндра и поршня постоянно появляются пузырьки воздуха, нарушающие целостность водного столба. Происходит обрыв этого столба при подъеме на высоту более 10 м.
Для того чтобы поднять воду на высоту 100 м, необходимо наличие сосущих сил в кроне дерева порядка 30 – 35 атм: на преодоление силы тяжести – 10 атм, сопротивления фильтрации через поперечные стенки сосудов – 20 – 25 атм. В природной обстановке леса такие величины часто регистрируются экспериментально. Поэтому с чисто физической точки зрения представляется возможным объяснить подъем воды на высоту 100 м и более.
В процесс е транспирации в листьях деревьев возникают сосущие силы, достигающие десятков атмосфер. Листья насасывают воду из стебля, вследствие чего в сосудах возникает отрицательное давление разрежение. Такое состояние можно наблюдать при помощи несложных приборов: в теплый летний день при интенсивной потере воды стволы деревьев уменьшаются в диаметре. Другой способ заключается в том, что при срезании интенсивно транспирирующей ветки в подкрашенной воде отмечается мгновенное проникновение краски через поверхность среза благодаря расширению сосудов.
В зависимости от анатомического строения древесины линейная скорость восходящего тока колеблется от 1 – 6 м/ч у хвойных и рассеянно-сосудистых древесных пород до 25 – 60 м/ч у кольцесосудистых. Такая скорость зарегистрирована летом в полдень. Скорость передвижения воды по дереву в течение суток изменяется и в основном соответствует интенсивности транспирации. Существует и светозависимый восходящий водный поток в растениях, тесно не связанный с транспирационной активностью (В.Г. Реуцкий).
В самом дереве быстрее всего вода передвигается в стволе и медленнее – в наиболее молодых ветвях. Среднее положение по этому показателю занимают старые ветви.
Особенности водного тока по стволу дерева:
· С помощью изотопной техники и введения в ствол красок было показано, что у большинства древесных растений водный ток в стволе передвигается по спирали. Это тесно связано с макростроением древесного ствола, что затрудняет ответ на вопрос, какая часть корневой системы питает водой ту или иную сторону кроны дерева.
· Передвижение воды в радиальном направлении осуществляется медленнее и происходит через поры на стенках сосудов и трахеид. Оно имеет значение в поддержании нормальной оводненности живых элементов древесины и коры.
· Неравномерность водного тока. Далеко не вся древесина служит местом проведения воды. У ядровых древесных растений (сосна, дуб) для этого служит только заболонь. При этом более активны в проведении воды последние годичные слои древесины. Это объясняется тем, что только эти слои переходят в древесину однолетних ветвей, тесно связанную с водопроводящей системой листьев. У хвойных, в частности у ели, проводящие пучки хвои сообщаются, по-видимому, с несколькими годичными кольцами. Старые годичные кольца просто не достигают кроны, они выклиниваются по мере увеличения высоты дерева. У ряда древесных пород (акация белая, фисташка, ясень) вода проводится всего лишь 1 – 3 последними годичными слоями заболони. Такая же картина наблюдается и у заболонных, и у спелодревесных пород (осина, береза, липа), но у них число годичных слоев, проводящих воду, несколько больше.
· Можно назвать и такую особенность восходящего тока в стволе дерева, как его изолированность. В общих чертах она присуща и травянистым растениям. Тем не менее, у древесных растений она выражена в большей степени. Проводящая воду древесина ствола отделена от внешней среды не только живыми клетками камбия и флоэмы, но и толстой пробкой или коркой, ограничивающих связь древесины с воздухом атмосферы.
· Такое свойство восходящего тока, как обратuмость, иногда наблюдается в природе у ряда древесных пород, в том числе у некоторых хвойных (ели, пихты), при укоренении нижних ветвей, соприкасающихся с почвой, или верхушек у вываленных деревьев. Ток воды здесь идет в обратном направлении – от морфологически верхнего конца к морфологически нижнему концу.
Восходящий ток обеспечивает все живые клетки растения водой и минеральными элементами. Очень важна роль восходящего тока и в обеспечении живых клеток корней, ствола и ветвей кислородом, ибо проницаемость тканей коры, камбия, древесины для газов весьма низка. Этот растворенный в воде восходящего тока кислород идет на осуществление процесса дыхания живых элементов дерева.
Восходящий ток влияет и на оводненность тканей, в частности древесины ствола. В молодом возрасте до образования ядра наибольшей влажностью обладает древесина хвойных пород. Это различие, очевидно, связано с отсутствием у них по сравнению с лиственными породами либриформа.
Содержание воды в древесине у большинства древесных растений увеличивается от внутренней части ствола к внешней и от основания ствола к его вершине. В пределах же кроны дерева количество воды в древесине увеличивается от вершины к основанию.
Резкие изменения оводненности древесины наблюдаются течение года. Так, у хвойных древесных растений самая низкая влажность отмечается в летние месяцы, а самая высокая – зимой. В поздневесеннее и раннеосеннее время влажность древесины занимает среднее положение. Влажность ядровой древесины практически остается неизменной и самой низкой. У лиственных древесных пород отмечено два периода пониженной влажности – летний и во второй половине зимы и два повышенной – весенний во время сокодвижения и зимний – в первой половине зимы.
Влажность древесины молодых деревьев несколько выше, а амплитуда колебаний ее больше, чем у старых деревьев. Содержание воды в древесине меняется и в течение летних суток: наиболее высокое рано утром, а низкое – в полдень.
Вода, запасенная в середине ствола, способна передвигаться в молодые побеги, что особенно важно при отсутствии доступной для растений воды в почве (засуха, морозы). Например, 100- летняя сосна может переносить засушливый период за счет внутренних запасов воды в стволе в течение целого месяца.
Перетекающая по растению вода отличается от метаболической воды,, которая непосредственно используется в различных процессах обмена веществ. Вода постоянно обменивается в клетках растений. С помощью современной техники, в частности изотопной, удалось показать, что самый быстрый обмен внутритканевой воды на внешнюю воду происходит в корнях растений, а самый медленный – в стеблях. Промежуточное положение занимают листья.
Дата добавления: 2016-11-18; просмотров: 2564 | Нарушение авторских прав | Изречения для студентов
Читайте также:
Рекомендуемый контект:
Поиск на сайте:
© 2015-2020 lektsii.org – Контакты – Последнее добавление
Источник