
Почечные канальцы и сосуды

Оглавление темы “Анатомия почки.”:
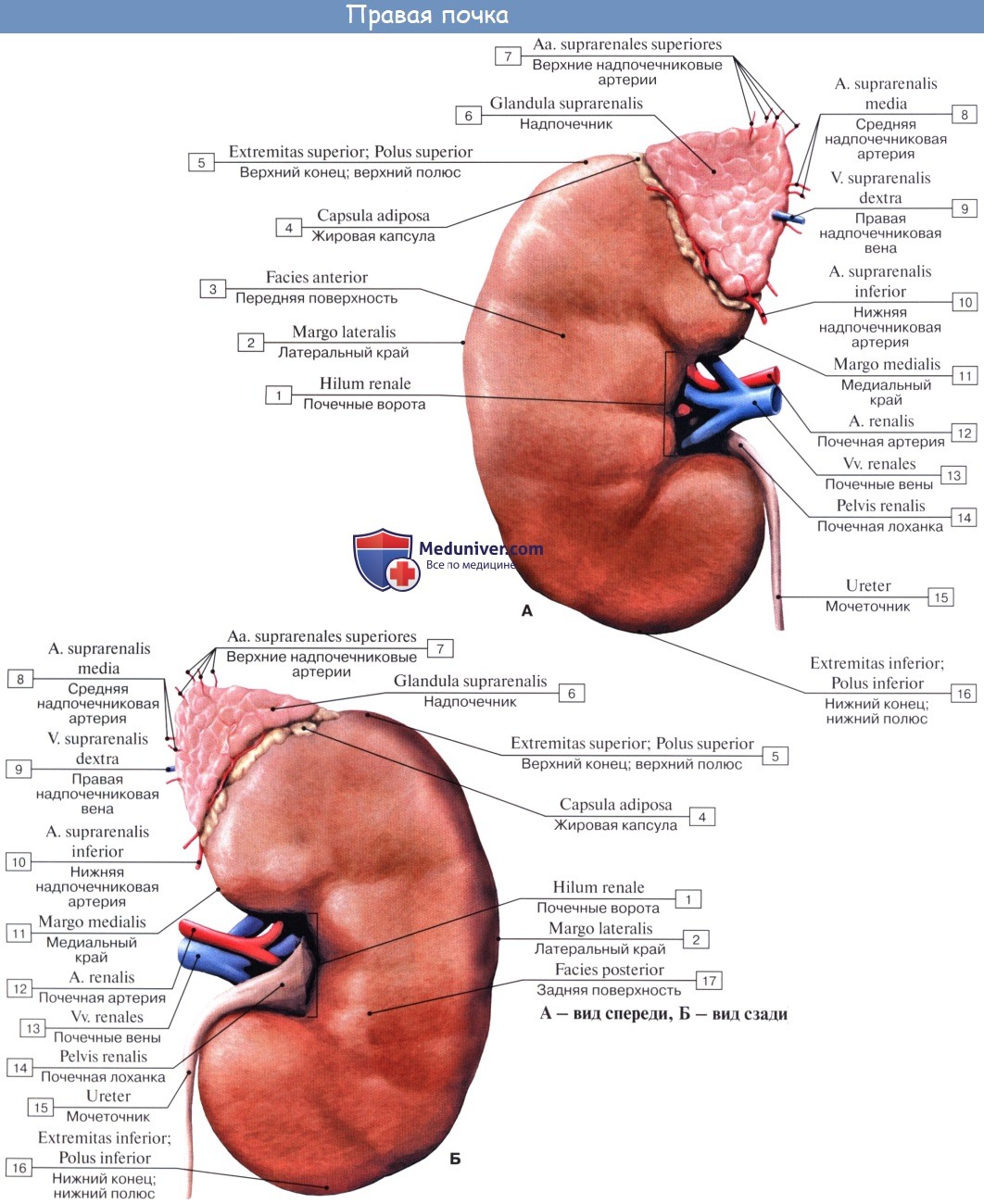
1. Почка, ren.
2. Топография почек. Оболочки почки. Фиксация почки.
3. Строение почек ( почки ). Кровоснабжение почек. Сосуды почек ( почки ).
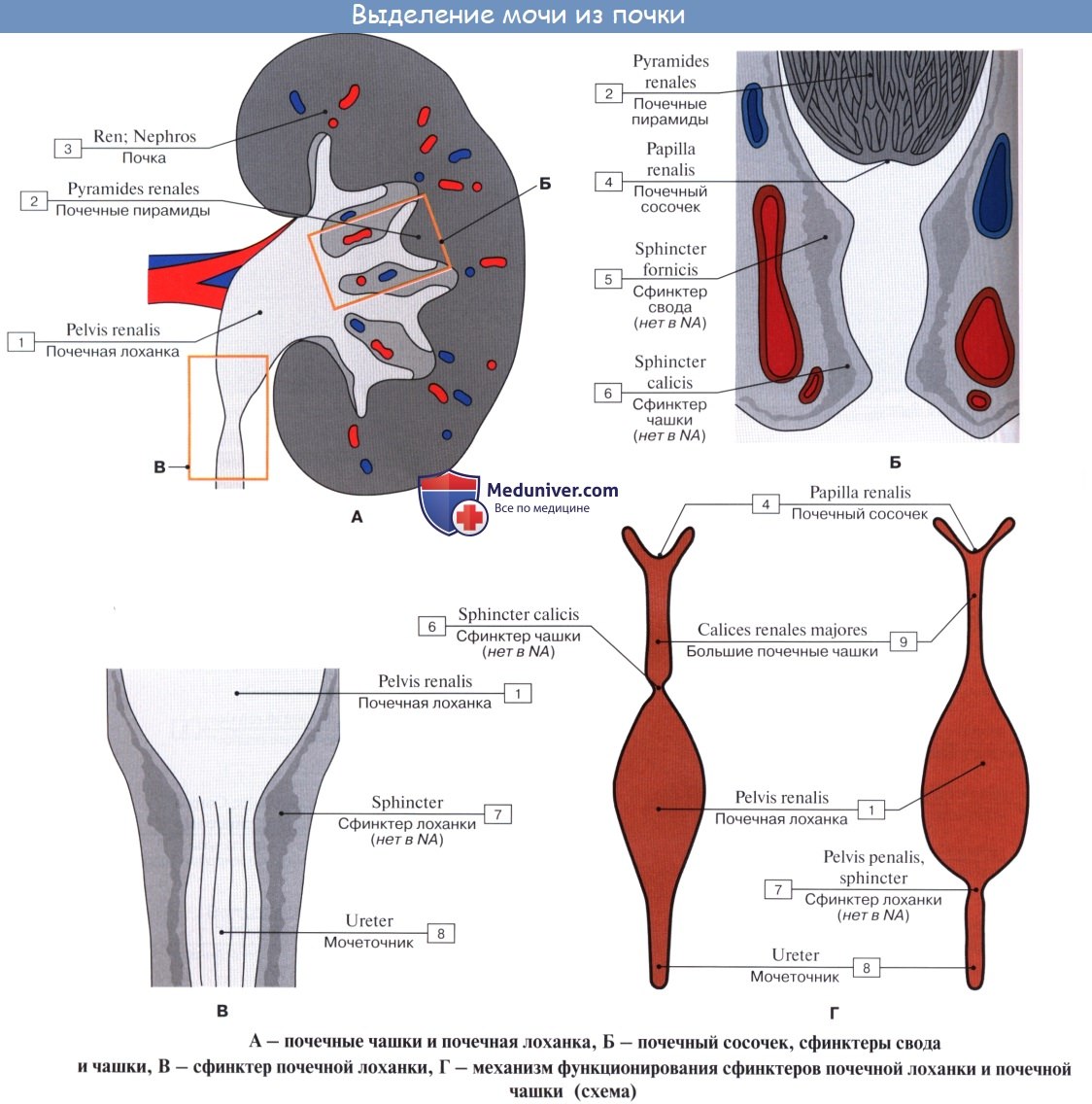
4. Почечная лоханка. Почечные чашки.
5. Рентгеноанатомия почки.
6. Сегментарное строение почки. Сегменты почки.
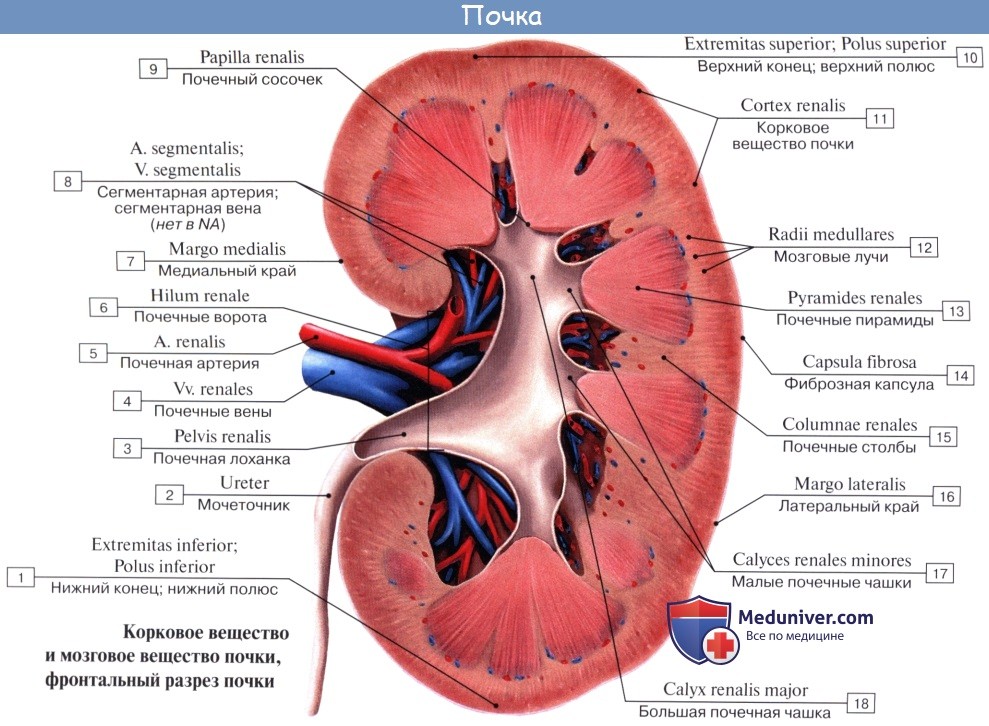
На продольном разрезе, проведенном через почку, видно, что почка в целом слагается, во-первых, из полости, sinus renalis, в которой расположены почечные чашки и верхняя часть лоханки, и, во-вторых, из собственно почечного вещества, прилегающего к синусу со всех сторон, за исключением ворот. В почке различают корковое вещество, cortex renis, и мозговое вещество, medulla renis.
Корковое вещество занимает периферический слой органа, имеет толщину около 4 мм. Мозговое вещество слагается из образований конической формы, носящих название почечных пирамид, pyramides renales. Широкими основаниями пирамиды обращены к поверхности органа, а верхушками—в сторону синуса.
Верхушки соединяются по две или более в закругленные возвышения, носящие название сосочков, papillae renales; реже одной верхушке соответствует отдельный сосочек. Всего сосочков имеется в среднем около 12.
Каждый сосочек усеян маленькими отверстиями, foramina papillaria; через foramina papillaria моча выделяется в начальные части мочевых путей (чашки). Корковое вещество проникает между пирамидами, отделяя их друг от друга; эти части коркового вещества носят название columnae renales. Благодаря расположенным в них в прямом направлении мочевым канальцам и сосудам пирамиды имеют полосатый вид. Наличие пирамид отражает дольчатое строение почки, характерное для большинства животных.
У новорожденного сохраняются следы бывшего разделения даже на наружной поверхности, на которой заметны борозды (дольчатая почка плода и новорожденного). У взрослого почка становится гладкой снаружи, но внутри, хотя несколько пирамид сливаются в один сосочек (чем объясняется меньшее число сосочков, нежели число пирамид), остается разделенной на дольки — пирамиды.
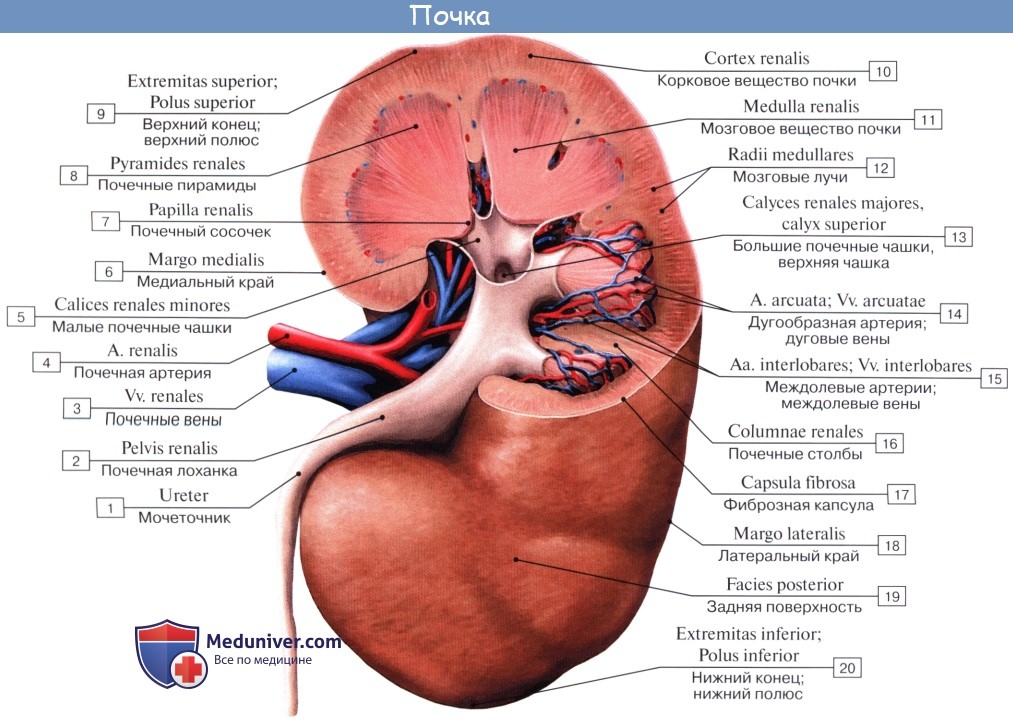
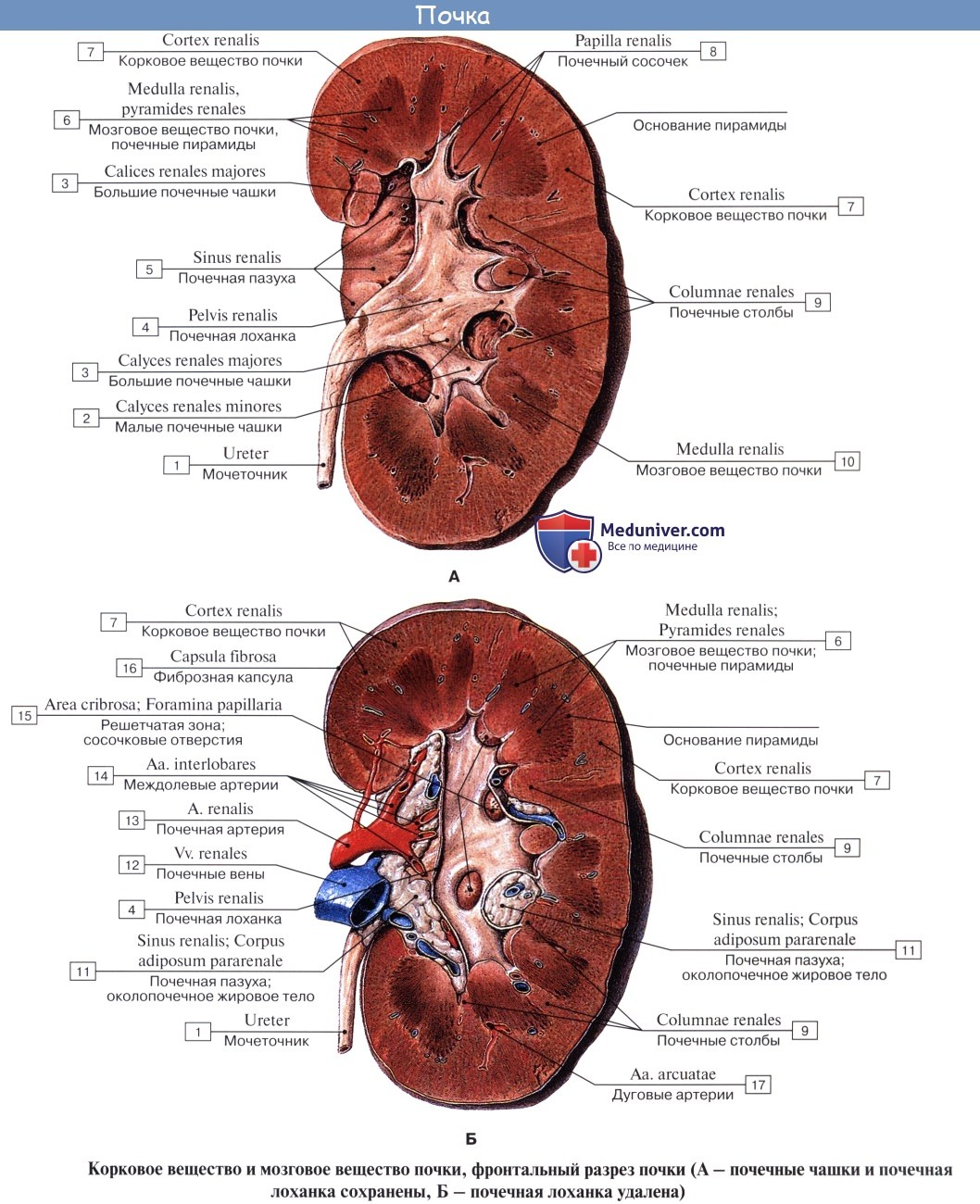
Полоски медуллярного вещества продолжаются также и в корковое вещество, хотя они заметны здесь менее отчетливо; они составляют pars radiata коркового вещества, промежутки же между ними — pars convoluta (convolutum — сверток).
Pars radiata и pars convoluta объединяют под названием lobulus corticalis.
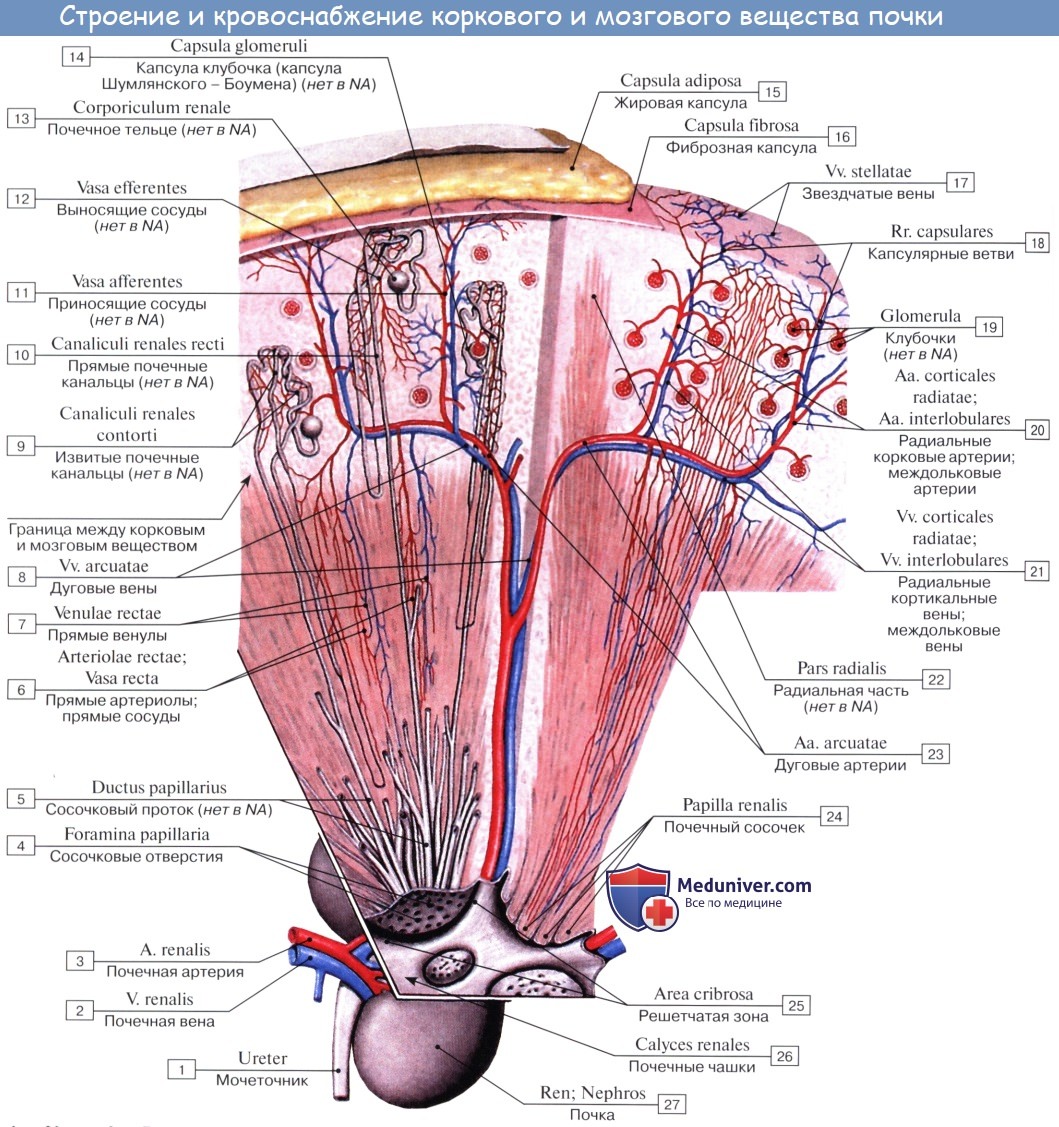
Почка представляет собой сложный экскреторный (выделительный) орган. Он содержит трубочки, которые называются почечными канальцами, tubuli renales. Слепые концы этих трубочек в виде двустенной капсулы охватывают клубочки кровеносных капилляров.
Каждый клубочек, glomerulus, лежит в глубокой чашеобразной капсуле, capsula glomeruli; промежуток между двумя листками капсулы составляет полость этой последней, являясь началом мочевого канальца. Glomerulus вместе с охватывающей его капсулой составляет почечное тельце, corpusculum renis.
Почечные тельца расположены в pars convoluta коркового вещества, где они могут быть видимы невооруженным глазом в виде красных точек. От почечного тельца отходит извитой каналец — tubulus renalis contdrtus, который находится уже в pars radiata коркового вещества. Затем каналец спускается в пирамиду, поворачивает там обратно, делая петлю нефрона, и возвращается в корковое вещество.
Конечная часть почечного канальца — вставочный отдел — впадает в собирательную трубочку, которая принимает несколько канальцев и идет по прямому направлению {tubulus renalis rectus) через pars radiata коркового вещества и через пирамиду. Прямые трубочки постепенно сливаются друг с другом и в виде 15 — 20 коротких протоков, ductus papillares, открываются foramina papillaria в области area cribrosa на вершине сосочка.
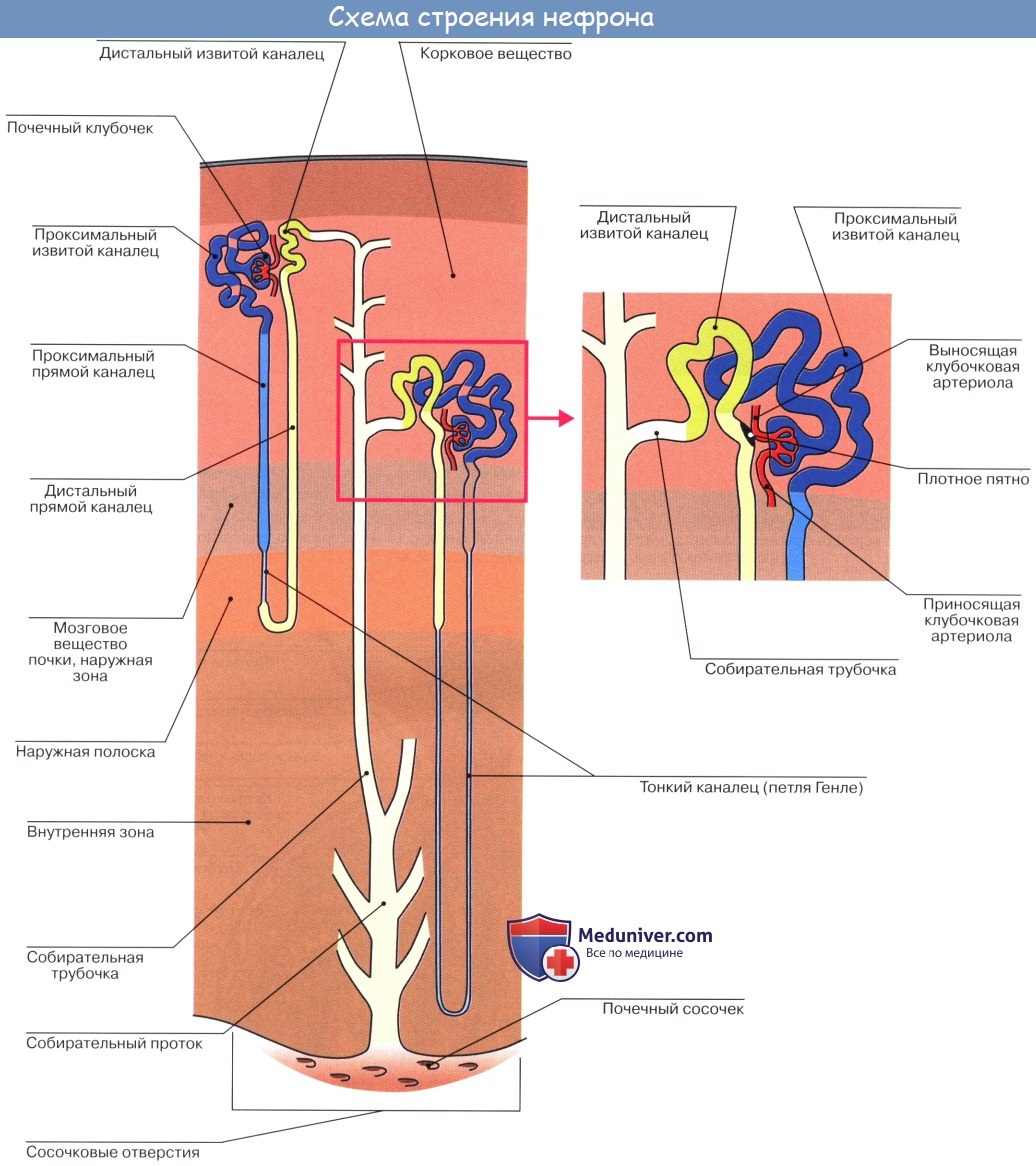
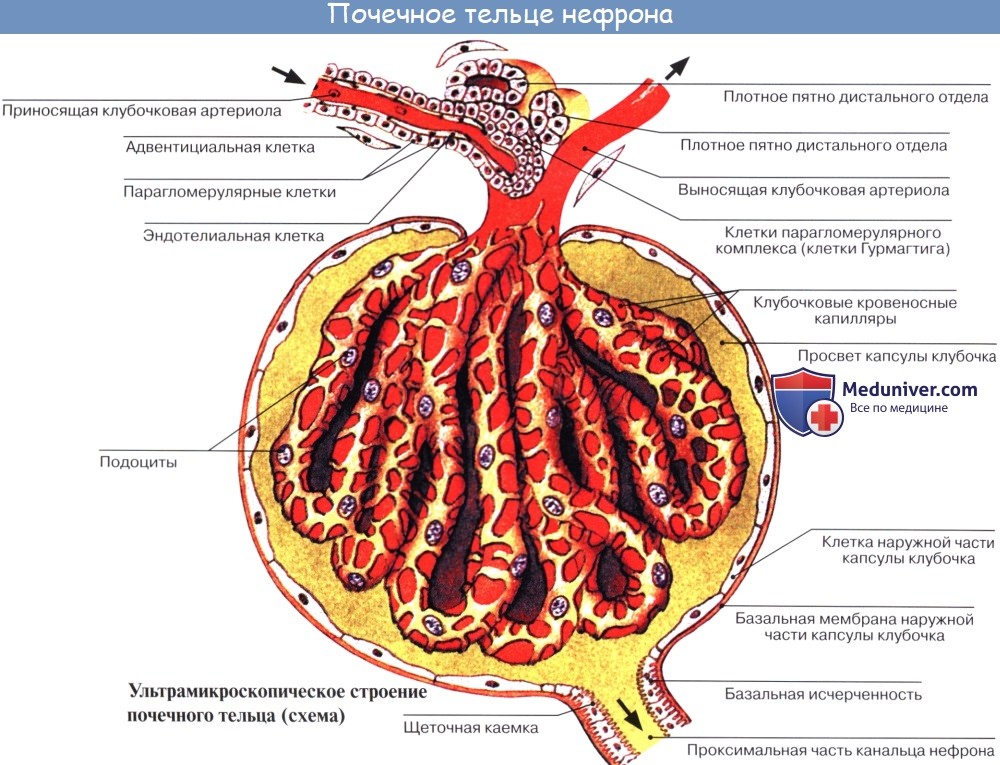
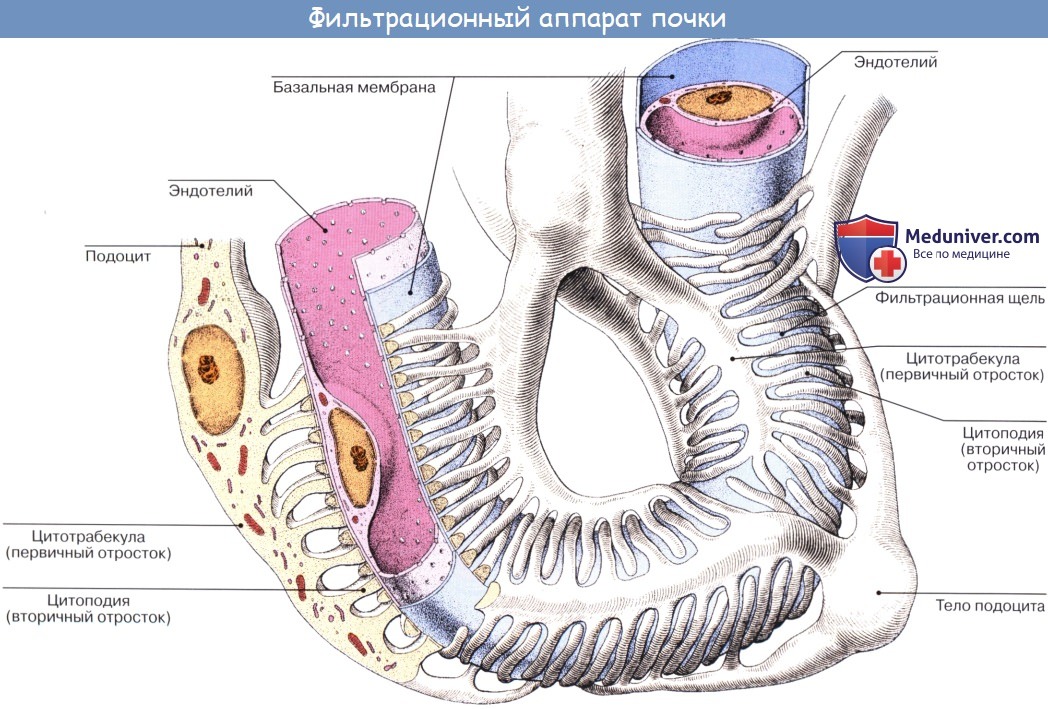
Почечное тельце и относящиеся к нему канальцы составляют структурно-функциональную единицу почки — нефрон, nephron. В нефроне образуется моча. Этот процесс совершается в два этапа: в почечном тельце из капиллярного клубочка в полость капсулы фильтруется жидкая часть крови, составляя первичную мочу, а в почечных канальцах происходит реабсорб-ция — всасывание большей части воды, глюкозы, аминокислот и некоторых солей, в результате чего образуется окончательная моча.
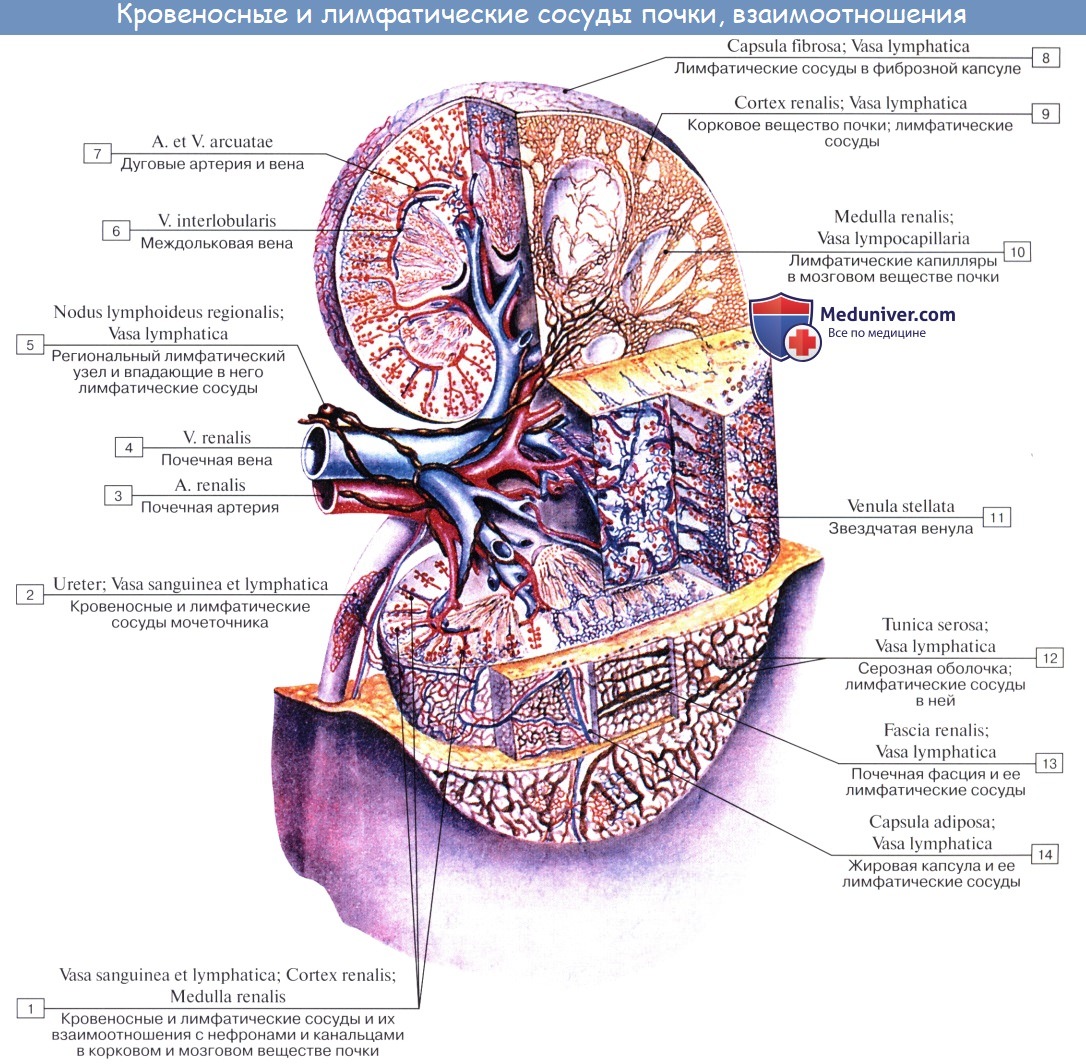
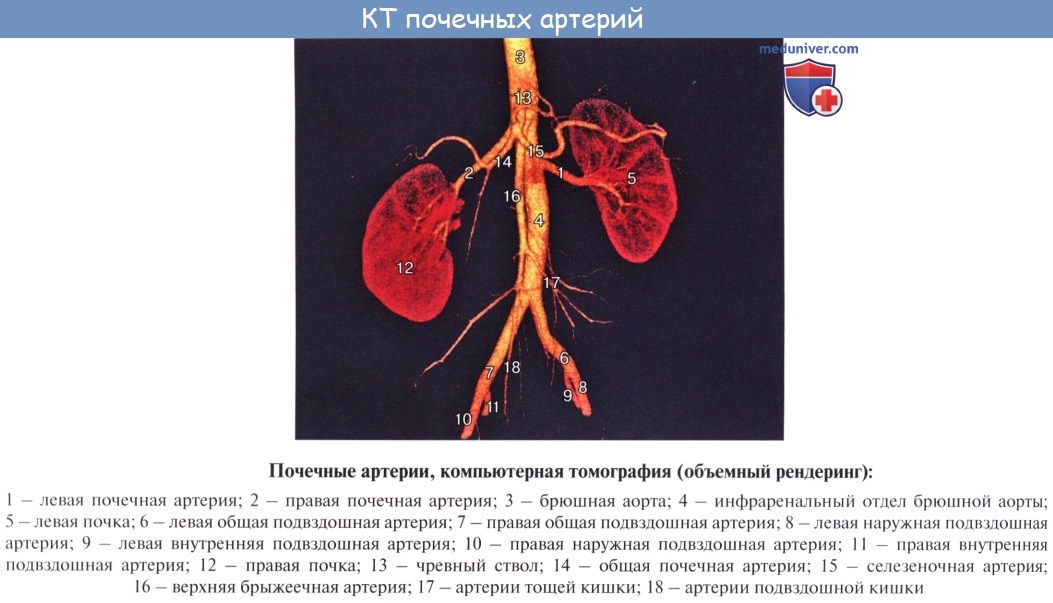
В каждой почке находится до миллиона нефронов, совокупность которых составляет главную массу почечного вещества. Для понимания строения почки и ее нефрона надо иметь в виду ее кровеносную систему. Почечная артерия берет начало от аорты и имеет весьма значительный калибр, что соответствует мочеотделительной функции органа, связанной с «фильтрацией» крови.
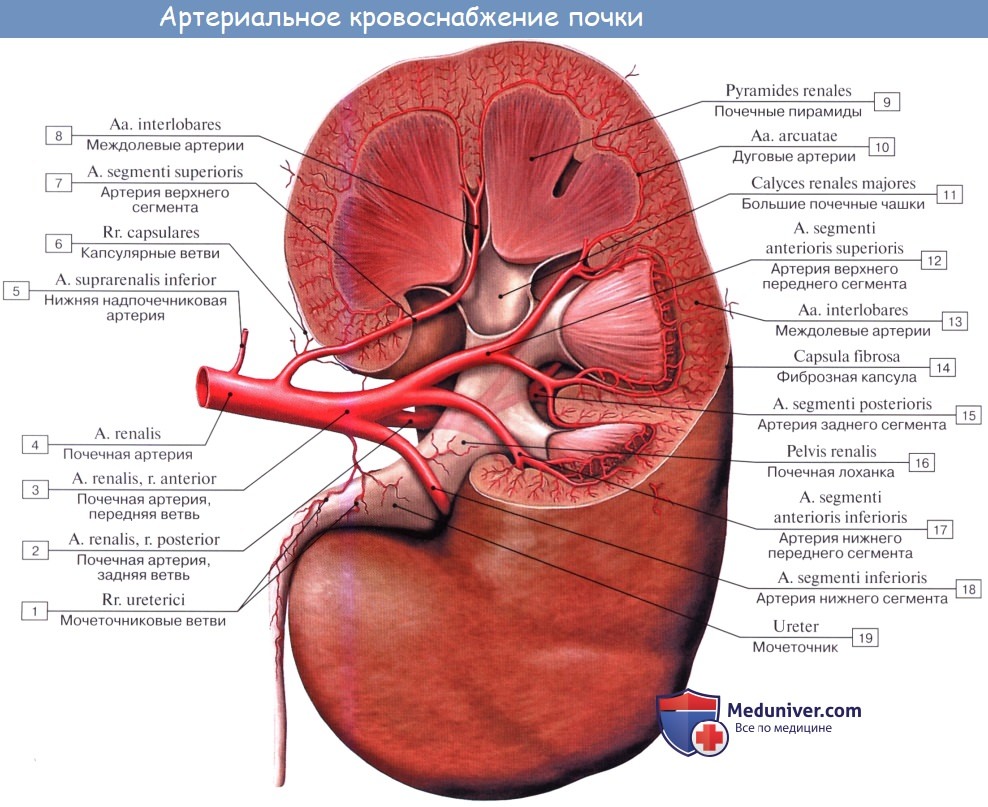
У ворот почки почечная артерия делится соответственно отделам почки на артерии для верхнего полюса, аа. polares superiores, для нижнего, аа. polares inferiores, и для центральной части почек, аа. centrales. В паренхиме почки эти артерии идут между пирамидами, т. е. между долями почки, и потому называются аа. interlobares renis. У основания пирамид на границе мозгового и коркового вещества они образуют дуги, аа. arcuatae, от которых отходят в толщу коркового вещества аа. interlobulares.
От каждой a. interlobularis отходит приносящий сосуд vas afferens, который распадается на клубок извитых капилляров, glomerulus, охваченный началом почечного канальца, капсулой клубочка. Выходящая из клубочка выносящая артерия, vas efferens, вторично распадается на капилляры, которые оплетают почечные канальцы и лишь затем переходят в вены. Последние сопровождают одноименные артерии и выходят из ворот почки одиночным стволом, v. renalis, впадающим в v. cava inferior.
Венозная кровь из коркового вещества оттекает сначала в звездчатые вены, venulae stellatae, затем в vv. interlobulares, сопровождающие одноименные артерии, и в vv. arcuatae. Из мозгового вещества выходят venulae rectae. Из крупных притоков v. renalis складывается ствол почечной вены. В области sinus renalis вены располагаются спереди от артерий.
Таким образом, в почке содержатся две системы капилляров; одна соединяет артерии с венами, другая — специального характера, в виде сосудистого клубочка, в котором кровь отделена от полости капсулы только двумя слоями плоских клеток: эндотелием капилляров и эпителием капсулы. Это создает благоприятные условия для выделения из крови воды и продуктов обмена.
Учебное видео анатомии почки
Другие видео уроки по данной теме находятся: Здесь.
– Также рекомендуем “Почечная лоханка. Почечные чашки.”
Редактор: Искандер Милевски. Дата последнего обновления публикации: 31.8.2020
Источник
Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 24 января 2017; проверки требуют 28 правок.
Нефрон (от греческого νεφρός (нефрос) — «почка») — структурно-функциональная единица почки. Нефрон состоит из почечного тельца, где происходит фильтрация, и системы канальцев, в которых осуществляются реабсорбция (обратное всасывание) и секреция веществ.
Структура и функции нефрона[править | править код]
Почечное тельце[править | править код]
1. Базальная мембрана
2. Капсула Боумена — Шумлянского — париетальная пластинка
3. Капсула Боумена — Шумлянского — висцеральная пластинка
3a. Подии (ножки) подоцита
3b. Подоцит
4. Пространство Боумена — Шумлянского
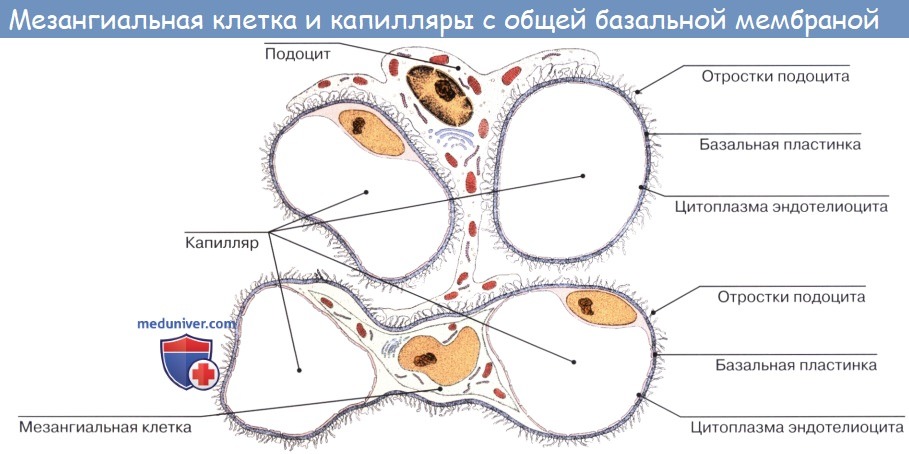
5a. Мезангий — Интрагломерулярные клетки
5b. Мезангий — Экстрагломерулярные клетки
6. Гранулярные (юкстагломерулярные) клетки
7. Плотное пятно
8. Миоцит (гладкая мускулатура)
9. Приносящая артериола
10. Клубочковые капилляры
11. Выносящая артериола
Нефрон начинается с почечного тельца, которое состоит из клубочка и капсулы Боумена — Шумлянского. Здесь осуществляется ультрафильтрация плазмы крови, которая приводит к образованию первичной мочи.
Типы нефронов[править | править код]
Различают три типа нефронов — интракортикальные нефроны (~85 %), юкстамедуллярные нефроны (~15 %) и субкапсулярные (суперфициальные).
- Почечное тельце интракортикального нефрона расположено в наружной части коркового вещества (внешняя кора) почки. Петля Генле у большинства интракортикальных нефронов имеет небольшую длину и располагается в пределах внешнего мозгового вещества почки.
- Почечное тельце юкстамедуллярного нефрона расположено в юкстамедуллярной коре, около границы коры почки с мозговым веществом. Большинство юкстамедуллярных нефронов имеют длинную петлю Генле. Их петля Генле проникает глубоко в мозговое вещество и иногда достигает верхушек пирамид
- Субкапсулярные (суперфициальные) находятся под капсулой.
Клубочек[править | править код]
Клубочек представляет собой группу сильно фенестрированных (окончатых) капилляров, получающих кровоснабжение от афферентной артериолы. Их также называют волшебной сетью (лат. rete mirabilis), так как газовый состав крови, проходящей через них, на выходе изменен незначительно (эти капилляры непосредственно не предназначены для газообмена). Гидростатическое давление крови создаёт движущую силу для фильтрации жидкости и растворённых веществ в просвет капсулы Боумена — Шумлянского.
Непрофильтровавшаяся часть крови из клубочков поступает в эфферентную артериолу. Эфферентная артериола поверхностно расположенных клубочков распадается на вторичную сеть капилляров, оплетающих извитые канальцы почек, эфферентные артериолы от глубоко расположенных (юкстамедуллярных) нефронов продолжаются в нисходящие прямые сосуды (лат. vasa recta), опускающиеся в мозговое вещество почек. Вещества, реабсорбированные в канальцах, в дальнейшем поступают в эти капиллярные сосуды.
Капсула нефрона[править | править код]
Капсула Боумена — Шумлянского окружает клубочек и состоит из висцерального (внутреннего) и париетального (внешнего) листков. Внешний листок представляет собой обычный однослойный плоский эпителий. Внутренний листок составлен из подоцитов, которые лежат на базальной мембране эндотелия капилляров, и ножки которых покрывают поверхность капилляров клубочка. Ножки соседних подоцитов образуют на поверхности капилляра интердигиталии. Промежутки между клетками в этих интердигиталиях и образуют, собственно, щели фильтра, затянутые мембраной. Размер этих фильтрационных пор ограничивает перенос крупных молекул и клеточных элементов крови.
Между внутренним листком капсулы и внешним, представленным простым, непроницаемым, плоским эпителием, лежит пространство, в которое поступает жидкость, профильтровавшаяся через фильтр, который сформирован мембраной щелей в интердигиталиях, базальной пластинкой капилляров и гликокаликсом, секретируемым подоцитами.
Нормальная скорость клубочковой фильтрации (СКФ) составляет 180—200 литров в сутки, что в 15—20 раз превышает объём циркулирующей крови — иными словами, вся жидкость крови за сутки успевает профильтроваться приблизительно двадцать раз.
Измерение СКФ является важной диагностической процедурой, её снижение может быть показателем почечной недостаточности.
Небольшие молекулы — такие, как вода, ионы Na+, Cl-, аминокислоты, глюкоза, мочевина, одинаково свободно проходят через клубочковый фильтр, так же проходят через него белки массой до 30 кДа, хотя, поскольку белки в растворе обычно несут отрицательный заряд, для них определённое препятствие составляет отрицательно заряженный гликокаликс. Для клеток и более крупных белков клубочковый ультрафильтр представляет непреодолимое препятствие. В результате, в пространство Боумена — Шумлянского, и далее в проксимальный извитой каналец, поступает жидкость, по составу отличающаяся от плазмы крови только отсутствием крупных белковых молекул.
Почечные канальцы[править | править код]
Проксимальный каналец[править | править код]
Микрофотография нефрона
1 — Клубочек (гломерула)
2 — Проксимальный каналец
3 — Дистальный каналец
Проксимальный каналец — наиболее длинная и широкая часть нефрона, проводящая фильтрат из капсулы Шумлянского — Боумена в петлю Генле.
Строение проксимального канальца[править | править код]
Проксимальный каналец построен из высокого цилиндрического эпителия с сильно выраженными микроворсинками апикальной мембраны (так называемая «щеточная кайма») и интердигитациями базолатеральной мембраны. Как микроворсинки, так и интердигитации значительно увеличивают поверхность клеточных мембран, усиливая тем самым их резорбтивную функцию.
Цитоплазма клеток проксимального канальца насыщена митохондриями, которые в большей степени находятся на базальной стороне клеток, тем самым обеспечивая клетки энергией, необходимой для активного транспорта веществ из проксимального канальца.
Транспортные процессы[править | править код]
| Реабсорбция |
|---|
| Na+: трансцеллюлярно (Na+ / K+-АТФаза, совместно с глюкозой — симпорт; Na+/Н+-обмен — антипорт), межклеточно |
| Cl-, K+, Ca2+, Mg2+: межклеточно |
| НСО3-: Н+ + НСО3- = СО2 (диффузия) + Н2О |
| Вода: осмос |
| Фосфат (регуляция ПТГ), глюкоза, аминокислоты, мочевые кислоты (симпорт с Na+) |
| Пептиды: расщепление до аминокислот |
| Белки: эндоцитоз |
| Мочевина: диффузия |
| Секреция |
| Н+: обмен Na+/H+, H+-АТФаза |
| NH3, NH4+ |
| Органические кислоты и основания |
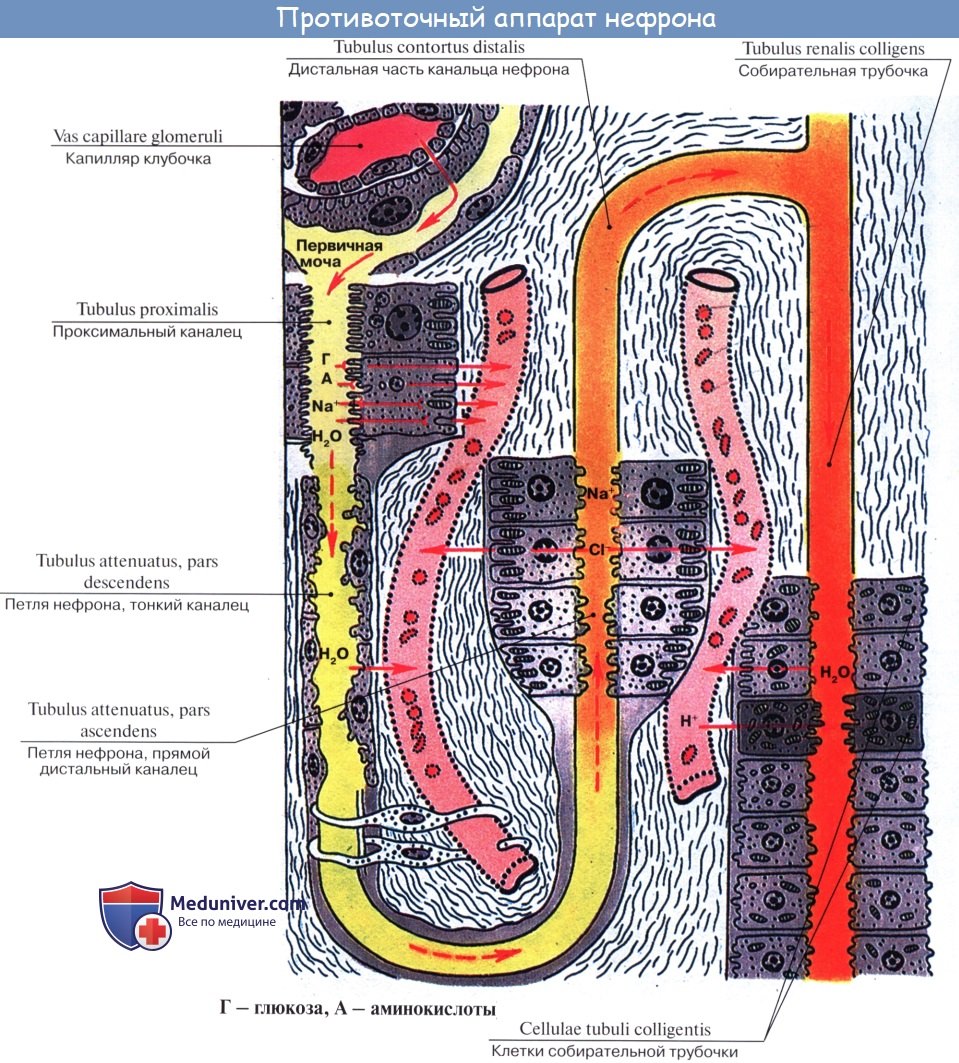
Петля Генле[править | править код]
Петля Генле — часть нефрона, соединяющая проксимальный и дистальный канальцы. Она имеет шпилечный изгиб в мозговом слое почки. Главная функция петли Генле состоит не в реабсорбции воды, а в сохранении ионов в мочевине по противоточному механизму в мозговом слое почки. Петля названа в честь Фридриха Густава Якоба Генле, немецкого патологоанатома.
Нисходящее колено петли Генле[править | править код]
Проксимальный извитой каналец в корковом веществе переходит в нисходящее колено петли Генле, которое спускается в мозговое вещество почки, образует там шпилькообразный изгиб, и переходит в восходящее колено петли Генле.
Транспортные процессы[править | править код]
Транспорт веществ:
| Вещество | Проницаемость |
| Ионы | Низкая проницаемость, активный транспорт отсутствует. |
| Мочевина | Умеренная пассивная проницаемость. |
| Вода | Высокая проницаемость, обусловленная присутствием аквапорина 1 как в апикальной, так и в базолатеральной мембранах клеток. Высокая осмолярность интерстиция мозгового вещества в сочетании с высокой водной проницаемостью эпителия приводит к реабсорбции большого объёма воды в этом отделе нефрона благодаря осмосу. |
Вследствие этого в нисходящем отделе петли Генле осмоляльность мочи резко возрастает и может достигать 1400 мосм/кг.
Гистология[править | править код]
Благодаря отсутствию активного транспорта клетки в данном отделе могут иметь сравнительно небольшой объём. Вместе с тем эффективный пассивный перенос воды требует малого расстояния диффузии. Вследствие этого, нисходящий отдел петли Генле построен из низкого кубического эпителия.
От кровеносных сосудов его можно отличить по отсутствию эритроцитов, а от толстых восходящих сегментов — по высоте эпителия.
Восходящее колено петли Генле[править | править код]
Транспортные процессы[править | править код]
| Тонкая восходящая часть | Реабсорбция NaCl (пассивно) |
| Толстая восходящая часть | Реабсорбция: NaCl (симпорт Na+/2Cl-/K+; Na+/K+-АТФаза + Cl–каналы) K+ (межклеточно) Ca2+, Mg2+ (регуляция ПТГ) NH4+ (симпорт Na+/2Cl-/NH4+) |
Дистальный извитой каналец[править | править код]
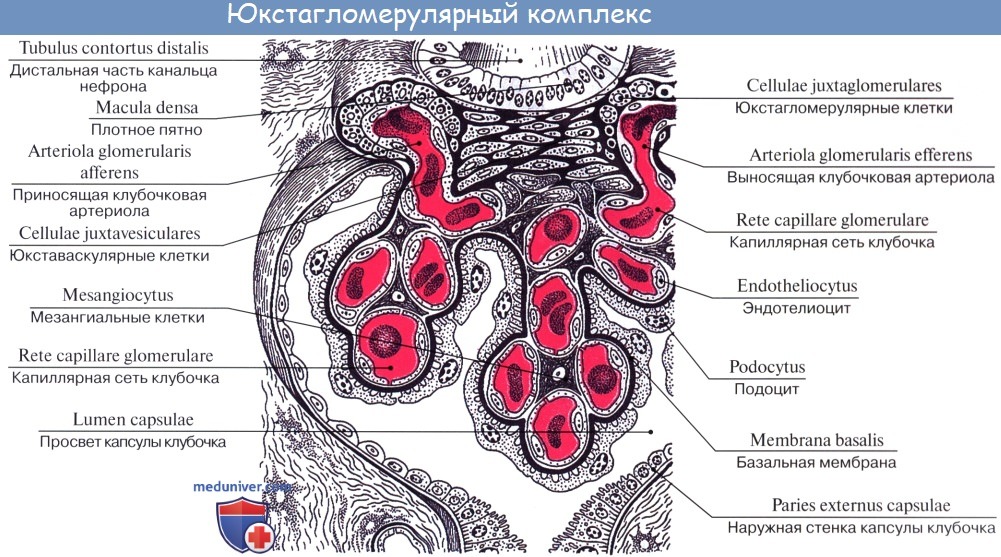
Юкстагломерулярный комплекс[править | править код]
Расположен в околоклубочковой зоне между приносящей и выносящей артериолами и состоит из трех основных частей:
Юкстагломерулярный аппарат участвует в синтезе ренина, который играет важнейшую роль в ренин-ангиотензиновой системе.
Ссылки[править | править код]
- Жизнь вопреки Хронической Почечной Недостаточности. Сайт: А. Ю. Денисова
- Нефрон, почечные клубочки. Состав мочи
- Реабсорбция в петле нефрона
- Global Dialysis
- Общество диализных пациентов НЕФРОН
Источник
Канальцевую часть нефрона принято делить на четыре отдела:
1) главный (проксимальный);
2) тонкий сегмент петли Генле;
3) дистальный;
4) собирательные трубки [Bargmann W., 1978].
Главный (проксимальный) отдел состоит из извилистой и прямой частей. Клетки извитой части имеют более сложное строение, чем клетки других отделов нефрона. Это высокие (до 8 мкм) клетки со щеточной каемкой, внутриклеточными мембранами, большим числом правильно ориентированных митохондрий, хорошо развитыми пластинчатым комплексом и эндоплазматической сетью, лизосомами и другими ультраструктурами (рис. 1). В их цитоплазме много аминокислот, основных и кислых белков, полисахаридов и активных SH-групп, высокоактивных дегидрогеназ, диафораз, гидролаз [Серов В. В., Уфимцева А. Г., 1977; Jakobsen N., Jorgensen F. 1975].
![Схема ультраструктуры клеток канальцев различных отделов нефрона [Rhodin J.( 1958]. 1 — клетка извитой части главного отдела; 2 — клетка прямой части главного отдела; 3 — клетка тонкого сегмента петли Генле; 4 — клетка прямой (восходящей) части дистального отдела; 5 — клетка извитой части дистального отдела; 6 — "темная" клетка связующего отдела и собирательной трубки; 7 — «светлая» клетка связующего отдела и собирательной трубки.](https://medbe.ru/upload/medialibrary/3c1/%D0%A1%D0%BD%D0%B8%D0%BC%D0%BE%D0%BA.JPG "Схема ультраструктуры клеток канальцев различных отделов нефрона [Rhodin J., 1958]. 1 — клетка извитой части главного отдела; 2 — клетка прямой части главного отдела; 3 — клетка тонкого сегмента петли Генле; 4 — клетка прямой (восходящей) части дистального отдела; 5 — клетка извитой части дистального отдела; 6 — \"темная\" клетка связующего отдела и собирательной трубки; 7 — «светлая» клетка связующего отдела и собирательной трубки.")
Рис. 1. Схема ультраструктуры клеток канальцев различных отделов нефрона [Rhodin J., 1958]. 1 — клетка извитой части главного отдела; 2 — клетка прямой части главного отдела; 3 — клетка тонкого сегмента петли Генле; 4 — клетка прямой (восходящей) части дистального отдела; 5 — клетка извитой части дистального отдела; 6 — “темная” клетка связующего отдела и собирательной трубки; 7 — «светлая» клетка связующего отдела и собирательной трубки.
Клетки прямой (нисходящей) части главного отдела в основном имеют то же строение, что и клетки извитой части, но пальцевидные выросты щеточной каемки более грубые и короткие, внутриклеточных мембран и митохондрий меньше, они не так строго ориентированы, значительно меньше цитоплазматических гранул.
Щеточная каемка состоит из многочисленных пальцевидных выростов цитоплазмы, покрытых клеточной мембраной и гликокаликсом. Их число на поверхности клетки достигает 6500, что увеличивает рабочую площадь каждой клетки в 40 раз [Bargmann W., 1978]. Эти сведения дают представление о поверхности, на которой совершается обмен в проксимальном отделе канальцев. В щеточной каемке доказана активность щелочной фосфатазы, АТФ-азы, 5-нуклеотидазы, аминопептидазы и ряда других ферментов [Wachsmuth Е., Stoye J. 1976]. Мембрана щеточной каемки содержит натрийзависимую транспортную систему. Считают, что гликокаликс, покрывающий микроворсинки щеточной каемки, проницаем для малых молекул. Большие молекулы поступают в каналец с помощью пиноцитоза, который осуществляется благодаря кратерообразным углублениям в щеточной каемке [Andrews P., Porter К., 1974].
Внутриклеточные мембраны образованы не только изгибами БМ клетки, но и латеральными мембранами соседних клеток, которые как бы перекрывают друг друга. Внутриклеточные мембраны являются по существу и межклеточными, что служит активному транспорту жидкости. При этом главное значение в транспорте придается базальному лабиринту, образованному выпячиваниями БМ внутрь клетки; он рассматривается как «единое диффузионное пространство» [Thoenes W., 1968].
Многочисленные митохондрии расположены в базальной части между внутриклеточными мембранами, что и создает впечатление их правильной ориентации. Каждая митохондрия, таким образом, заключена в камере, образованной складками внутри- и межклеточных мембран. Это позволяет продуктам энзиматических процессов, развивающихся в митохондриях, легко выходить за пределы клетки. Энергия, вырабатываемая в митохондриях, служит как транспорту вещества, так и секреции, осуществляемой с помощью гранулярной эндоплазматической сети и пластинчатого комплекса, который претерпевает циклические изменения в различные фазы диуреза.
Ультраструктура и ферментохимия клеток канальцев главного отдела объясняют его сложную и дифференцированную функцию. Щеточная каемка, как и лабиринт внутриклеточных мембран, является своеобразным приспособлением для колоссальной по объему функции реабсорбции, выполняемой этими клетками. Ферментная транспортная система щеточной каемки, зависимая от натрия, обеспечивает реабсорбцию глюкозы, аминокислот, фосфатов [Наточин Ю. В., 1974; Kinne R., 1976]. С внутриклеточными мембранами, особенно с базальным лабиринтом, связывают реабсорбцию воды, глюкозы, аминокислот, фосфатов и ряда других веществ [Kinne R., 1976], которую выполняет натрийнезависимая транспортная система мембран лабиринта.
Особый интерес представляет вопрос о канальцевой реабсорбции белка. Считают доказанным, что весь фильтрирующийся в клубочках белок реабсорбируется в проксимальном отделе канальцев, чем объясняется его отсутствие в моче здорового человека. Это положение основывается на многих исследованиях, выполненных, в частности, с помощью электронного микроскопа. Так, транспорт белка в клетке проксимального канальца изучен в опытах с микроинъекцией меченного ¹³¹I альбумина непосредственно в каналец крысы с последующей электронно-микроскопической радиографией этого канальца [Maunsbach А., 1966].
Альбумин находят прежде всего в инвагинатах мембраны щеточной каемки, затем в пиноцитозных пузырьках, которые сливаются в вакуоли. Белок с вакуолей появляется затем в лизосомах и пластинчатом комплексе (рис. 2) и расщепляется гидролитическими ферментами [Thoenes W., Langer К., 1969]. Вероятнее всего, «основные усилия» высокой дегидрогеназной, диафоразной и гидролазной активности в проксимальном отделе канальцев направлены на реабсорбцию белка.
![Схема реабсорбции белка клеткой канальцев главного отдела [Thoenes W., Langer К., 1969]. 1 — микропиноцитоз у основания щеточной каемки; Mvb —вакуоли, содержащие белок ферритин; II — заполненные ферритином вакуоли (а) перемещаются к базальной части клетки; б — лизосома; в — слияние лизосомы с вакуолью; г — лизосомы с инкорпорированным белком; АГ — пластинчатый комплекс с цистернами, содержащими КФ (окрашены в черный цвет); III — выделение через БМ низкомолекулярныхфрагментов реабсорбированного белка, образовавшихся после «переваривания» в лизосомах (показано двойными стрелками).](https://medbe.ru/upload/medialibrary/815/%D0%A1%D0%BD%D0%B8%D0%BC%D0%BE%D0%BA.JPG "Схема реабсорбции белка клеткой канальцев главного отдела [Thoenes W., Langer К., 1969]. I — микропиноцитоз у основания щеточной каемки; Mvb —вакуоли, содержащие белок ферритин; II — заполненные ферритином вакуоли (а) перемещаются к базальной части клетки; б — лизосома; в — слияние лизосомы с вакуолью; г — лизосомы с инкорпорированным белком; АГ — пластинчатый комплекс с цистернами, содержащими КФ (окрашены в черный цвет); III — выделение через БМ низкомолекулярных фрагментов реабсорбированного белка, образовавшихся после «переваривания» в лизосомах (показано двойными стрелками).")
Рис. 2. Схема реабсорбции белка клеткой канальцев главного отдела [Thoenes W., Langer К., 1969].
I — микропиноцитоз у основания щеточной каемки; Mvb —вакуоли, содержащие белок ферритин;
II — заполненные ферритином вакуоли (а) перемещаются к базальной части клетки; б — лизосома; в — слияние лизосомы с вакуолью; г — лизосомы с инкорпорированным белком; АГ — пластинчатый комплекс с цистернами, содержащими КФ (окрашены в черный цвет);
III — выделение через БМ низкомолекулярных фрагментов реабсорбированного белка, образовавшихся после «переваривания» в лизосомах (показано двойными стрелками).
В связи с этими данными становятся понятными механизмы “повреждения” канальцев главного отдела. При НС любого генеза, протеинурических состояниях изменения эпителия канальцев проксимального отдела в виде белковой дистрофии (гиалиново-капельной, вакуольной) отражают резорбционную недостаточность канальцев в условиях повышенной порозности гломерулярного фильтра для белка [Давыдовский И. В., 1958; Серов В. В., 1968]. Нет необходимости видеть в изменениях канальцев при НС первично-дистрофические процессы.
В равной мере нельзя рассматривать и протеинурию как результат только повышенной порозности гломерулярного фильтра. Протеинурия при нефрозах отражает как первичное повреждение фильтра почки, так и вторичное истощение (блокаду) ферментных систем канальцев, осуществляющих реабсорбцию белка.
При ряде инфекций и интоксикаций блокада ферментных систем клеток канальцев главного отдела может наступить остро, поскольку эти канальцы первыми подвергаются действию токсинов и ядов при их элиминации почками. Активация гидролаз лизосомного аппарата клетки завершает в ряде случаев дистрофический процесс развитием некроза клетки (острый нефроз). В свете приведенных данных становится понятной патология «выпадения» ферментов канальцев почек наследственного порядка (так называемые наследственные канальцевые ферментопатии). Определенная роль в повреждении канальцев (тубулолизис) отводится антителам, реагирующим с антигеном тубулярной базальной мембраны и щеточной каемки.
Клетки тонкого сегмента петли Генле характеризуются той особенностью, что внутриклеточные мембраны и пластинки пересекают тело клетки на всю ее высоту, образуя в цитоплазме щели шириной до 7 нм [Bargmann W., 1978]. Создается впечатление, что цитоплазма состоит из отдельных сегментов, причем часть сегментов одной клетки как бы вклинивается между сегментами соседней клетки. Ферментохимия тонкого сегмента отражает функциональную особенность этого отдела нефрона, который как дополнительное приспособление уменьшает до минимума фильтрационный заряд воды и обеспечивает ее «пассивную» резорбцию [Уфимцева А. Г., 1963].
Соподчиненная работа тонкого сегмента петли Генле, канальцев прямой части дистального отдела, собирательных трубок и прямых сосудов пирамид обеспечивает осмотическое концентрирование мочи на основе противоточного умножителя [Wirz Н. et al, 1951]. Новые представления о пространственной организации противоточно-множительной системы (рис. 3) убеждают в том, что концентрирующая деятельность почки обеспечивается не только структурно-функциональной специализацией различных отделов нефрона, но и высокоспециализированным взаиморасположением канальцевых структур и сосудов почки [Перов Ю. Л., 1975; Kriz W., Lever А., 1969].
![Схема расположения структур противоточно-множительной системы в мозговой веществе почки [Kriz W., Lever А., 1969]. 1 - артериальный прямой сосуд; 2 — венозный прямой сосуд; 3 — тонкий сегмент петли Генле; 4 — прямая часть дистального отдела; СТ — собирательные трубки; К—капилляры.](https://medbe.ru/upload/medialibrary/672/%D0%A1%D0%BD%D0%B8%D0%BC%D0%BE%D0%BA.JPG "Схема расположения структур противоточно-множительной системы в мозговой веществе почки [Kriz W., Lever А., 1969]. 1 - артериальный прямой сосуд; 2 — венозный прямой сосуд; 3 — тонкий сегмент петли Генле; 4 — прямая часть дистального отдела; СТ — собирательные трубки; К—капилляры.")
Рис. 3. Схема расположения структур противоточно-множительной системы в мозговой веществе почки [Kriz W., Lever А., 1969]. 1 – артериальный прямой сосуд; 2 — венозный прямой сосуд; 3 — тонкий сегмент петли Генле; 4 — прямая часть дистального отдела; СТ — собирательные трубки; К — капилляры.
Дистальный отдел канальцев состоит из прямой (восходящей) и извитой частей. Клетки дистального отдела ультраструктурно напоминают клетки проксимального отдела. Они богаты сигарообразными митохондриями, заполняющими пространства между внутриклеточными мембранами, а также цитоплазматическими вакуолями и гранулами вокруг ядра, расположенного апикально, но лишены щеточной каемки. Эпителий дистального отдела богат аминокислотами, основными и кислыми белками, РНК, полисахаридами и реактивными SH-группами; для него характерна высокая активность гидролитических, гликолитических ферментов и ферментов цикла Кребса.
Сложность устройства клеток дистальных канальцев, обилие митохондрий, внутриклеточных мембран и пластического материала, высокая ферментативная активность свидетельствуют о сложности их функции — факультативной реабсорбции, направленной на поддержание постоянства физико-химических условий внутренней среды. Факультативная реабсорбция регулируется в основном гормонами задней доли гипофиза, надпочечников и ЮГА почки.
Местом приложения действия антидиуретического гормона гипофиза (АДГ), в почке, «гистохимическим плацдармом» этой регуляции служит система гиалуроновая кислота — гиалуронидаза, заложенная в пирамидах, главным образом в их сосочках. Альдостерон, по некоторым данным, и кортизон влияют на уровень дистальной реабсорбции прямым включением в ферментную систему клетки, обеспечивающую перенос ионов натрия из просвета канальца в интерстиции почки. Особое значение в этом процессе принадлежит эпителию прямой части дистального отдела, причем дистальный эффект действия альдостерона опосредован секрецией ренина, закрепленной за клетками ЮГА. Ангиотензин, образующийся под действием ренина, не только стимулирует секрецию альдостерона, но и участвует в дистальной реабсорбции натрия.
В извитой части дистального отдела канальца, там, где он подходит к полюсу сосудистого клубочка, различают macula densa [Kaissling В. et al., 1977]. Эпителиальные клетки в этой части становятся цилиндрическими, их ядра — гиперхромными; они располагаются полисадообразно, причем непрерывной базальной мембраны здесь нет. Клетки macula densa имеют тесные контакты с гранулированными эпителиоидными клетками и lacis-клетками ЮГА, что обеспечивает влияние химического состава мочи дистального канальца на гломерулярный кровоток и, наоборот гормональные влияния ЮГА на macula densa.
Со структурно-функциональной особенностью канальцев дистального отдела, их повышенной чувствительностью к кислородному голоданию связано до некоторой степени их избирательное поражение при острых гемодинамических повреждениях почек, в патогенезе которых основную роль играют глубокие нарушения почечного кровообращения с развитием аноксии тубулярного аппарата. В условиях острой аноксии клетки дистальных канальцев подвергаются воздействию содержащей токсические продукты кислой мочи, что ведет к их поражению вплоть до некроза. При хронической аноксии клетки дистального канальца чаще, чем проксимального, подвергаются атрофии.
Собирательные трубки, выстланные кубическим, а в дистальных отделах цилиндрическим эпителием (светлые и темные клетки) с хорошо развитым базальным лабиринтом, высокопроникаемы для воды. С темными клетками связывают секрецию ионов водорода, в них обнаружена высокая активность карбоангидразы [Зуфаров К. А. и др., 1974]. Пассивный транспорт воды в собирательных трубках обеспечивается особенностями и функции противоточно-множительной системы [Wirz Н., 1953].
Заканчивая описание гистофизиологии нефрона, следует остановиться на его структурно-функциональном различии в разных отделах почки. На этом основании выделяют кортикальные и юкстамедуллярные нефроны, различающиеся строением клубочков и канальцев, а также своеобразием функции; различно и кровоснабжение этих нефронов.
Клиническая нефрология
под ред. Е.М. Тареева
Опубликовал Константин Моканов
Источник