
Покрытосеменных растений отсутствие сосудов

Определение сосудистых растений
Сосудистый растение является одним из ряда растений со специализированными сосудистая ткань, Два типа сосудистых ткань, ксилема а также флоэма, несут ответственность за перемещение воды, минералов и продуктов фотосинтез по всему заводу. В отличие от несосудистого растения, сосудистое растение может расти намного больше. Сосудистая ткань внутри обеспечивает транспортировку воды на большую высоту, позволяя сосудистому растению расти вверх, чтобы поймать солнце.
Структура сосудистых растений
Внутри сосудистого растения структура сильно отличается от структуры несосудистого растения. У несосудистых растений дифференциация между клетками практически отсутствует. У сосудистых растений специализированные сосудистые ткани располагаются по уникальным рисункам, в зависимости от деления и вид сосудистое растение принадлежит.
Ксилема, состоящая в основном из структурного белка лигнина и мертвых клеток, специализируется на транспортировке воды и минералов от корней к листьям. Сосудистое растение делает это, создавая давление на воду по нескольким направлениям. В корнях вода впитывается в ткани. Вода течет в ксилему и создает восходящее давление. На листьях вода используется и испаряется из устьице, Говорят, что эти маленькие поры проходят, что тянет вверх столб воды в ксилеме. Через действия адгезия и сплоченность, вода движется вверх через ксилему, как напиток через соломинку. Этот процесс можно увидеть ниже.

В листьях происходит фотосинтез. Сосудистое растение, как низшие растения и водоросли Используйте тот же процесс, чтобы извлечь энергию от солнца и сохранить ее в связях глюкозы. Этот сахар превращается в другие формы и должен транспортироваться в части растения с невозможностью фотосинтеза, такие как стебель и корни. Эта флора специально разработана для этой цели. В отличие от ксилемы, флоэма состоит из частично живых клеток, которые помогают облегчить транспорт сахаров через транспортные белки, найденные в клетка Мембраны. Эта флоэма также связана с ксилемой и может добавлять воду, чтобы помочь разбавить и переместить сахар. Коммерчески собранный, это известно как сок или сироп, такой как кленовый сироп.
Жизненный цикл сосудистого растения
Сосудистые растения, как и все растения, демонстрируют смена поколений, Это означает, что есть две формы растения, спорофит и гаметофит, Спорофит, а диплоид организм, проходит через мейоз производить гаплоидный Спор. Спора перерастает в новый организм – гаметофит. Гаметофит отвечает за образование гамет, способных слиться во время половое размножение.
Эти гаметы, сперма и яйцеклетка, сливаются вместе, образуя зигота, которое является новым поколением диплоидных спорофитов. У некоторых растений эта зигота будет развиваться непосредственно в новый организм. В других зигота развивается в семя, которое рассеяно и должно иметь период покоя или некоторый сигнал активации, чтобы начать расти. Сосудистое растение, которое ближе к мхам и несосудистым растениям, с большей вероятностью будет иметь независимые чередующиеся поколения. Посевные растения, как правило, имеют сильно сниженный гаметофит, который, как правило, полностью зависит от спорофита и живет в нем. Различие едва заметно между этими двумя организмами, кроме количества ДНК, которое они несут в своих клетках (гаплоид против диплоида) и процессов клеточного деления, которые они используют.
Классификация сосудистых растений
Сосудистые растения представляют собой эмбриофиты, которые являются большими клады или родственная группа, состоящая как из несосудистых, так и из сосудистых растений. Эмбриофиты далее разбиваются на бриофиты, включая мхов, печеночники и несосудистые растения, и Tracheophyta. Поскольку трахея у людей является проходом для воздуха, термин трахеофит относится к сосудистой ткани сосудистых растений.
Трахеофиты далее делятся на отделы. Подразделения отличаются главным образом тем, как функционируют их споры и гаметофиты. У папоротников и клубных мхов гаметофит становится свободно живущим поколением. У голосеменных (хвойных) и покрытосеменных (цветковых растений) гаметофит зависит от спорофита. Гаметы, развитые внутри, становятся семенем, формируя следующее поколение спорофитов. В то время как каждое сосудистое растение имеет чередование поколений с доминирующим спорофитом, они различаются по способу распределения спор и семян.
Примеры сосудистых растений
Ежегодный Vs. круглогодичный
Некоторые растения, однолетние растения, завершают свой жизненный цикл в течение одного года. Если бы вы купили однолетнее растение в магазине, посадили его в своем саду и собрали все сброшенные семена, растение не вернулось бы в следующем году. Ежегодно, как правило, травянистый Имеется в виду их стебли и корни и не очень структурированные и жесткие. Хотя растения могут стоять высокими, это в основном связано с тургор давление на клеточных стенках растения.
Многолетнее растение немного отличается. Хотя это может быть и травянистое растение, оно вернется на несколько лет, даже если вы соберете все семена. В течение зимы сосудистое растение способно хранить сахар в корнях и полностью избегать замерзания. Весной растение может возобновить рост и попытаться еще раз произвести потомство. Хотя методы размножения отражают миллионы лет эволюции, они не отражают сосудистые растения по сравнению с несосудистыми.
Монокот против двудольных
В пределах покрытосеменных растений или цветковых растений существует огромное деление. В то время как однодольные и двудольные растения являются сосудистыми растениями, они различаются по способу формирования их семян и способу их роста. В однодольные Выращивание происходит под почвой, так как отдельные листья начинаются от корней и растут вверх. Кукуруза является однодольным, как и многие виды трав, включая пшеницу и ячмень. На других посевных растениях, таких как бобы и горох, есть два листа семядолей, которые делают их двудольными. Сосудистую ткань однодольных можно увидеть справа на изображении ниже.

В двудольным точка роста находится над почвой, и это приводит к тому, что растения разветвляются в нескольких направлениях. Таким образом, сосудистая ткань в двудольном ветвь разветвлена, где в однодольном она идет параллельно. Обратите внимание, как сосудистая ткань у этих растений создает организованные пучки. Этот шаблон создает легкие возможности ветвления. Эти изменения в сосудистой ткани представляют различные способы формирования листьев для сбора света, наблюдаемого в двух типах сосудистых растений.
викторина
1. Что из перечисленного НЕ является сосудистым растением?A. Красное деревоB. Мох C. Мир Лилли
Ответ на вопрос № 1
В верно. Мох – это несосудистое растение, то есть оно не имеет дифференцированных сосудистых тканей. Мхи могут быть многолетними, поскольку они могут бездействовать в течение зимних месяцев, чтобы выжить. Они не могут, однако, расти очень высокими, потому что они ограничены в распределении и использовании воды.
2. Какова цель ксилемы в сосудистом растении?A. Ксилем несет сахар вокруг растенияB. Ксилем движет воду от корней к побегамC. Ксилем переносит продукты фотосинтеза
Ответ на вопрос № 2
В верно. Первый и третий ответы одинаковы, поскольку продукты фотосинтеза – это сахара. Флоема переносит эти продукты вокруг растения, в то время как ксилема перемещает воду из земли в и из листьев. Это обеспечивает воду, давление тургора и источник питательных веществ.
3. Клубные мхи – это уникальный организм. Как и мхи, они не создают семя и используют споры для размножения. В отличие от мхов, они имеют различимые ткани, которые транспортируют воду по всему растению. Клубничные мхи могут расти значительно выше обычного мха. Какие из следующих утверждений верно?A. Клаб-Мосс – это сосудистое растениеB. Клаб-Мосс – несосудистое растениеC. Клаб-мох не сосудистый и не сосудистый
Ответ на вопрос № 3
верно. Сосудистое растение, независимо от жизненного цикла, определяется различимыми тканями сосудов. Это позволяет булаву-мху перемещать воду на гораздо большую высоту, чем обычный мох, увеличивая их способность поглощать солнечный свет.
Ссылки
- Hartwell, L.H., Hood, L., Goldberg, M.L., Reynolds, A.E. & Silver, L.M. (2011). генетика : От генов к геномам Бостон: Макгроу Хилл.
- Kaiser, M.J., Attrill, M.J., Jennings, S., Thomas, D.N., Barnes, D.K., Brierley, A.S. & Hiddink, J.G. (2011). морской экология : Процессы, системы и воздействия. Нью-Йорк: издательство Оксфордского университета.
- McMahon, M.J., Kofranek, A.M. & Rubatzky, V.E. (2011). Наука о растениях: рост, развитие и использование культурных растений (5-е изд.). Бостон: Прентинс Холл.
Источник
Отдел покрытосеменные (цветковые) самый многочисленный, он включает 235-250 тысяч видов. Его представители обитают по всему миру:
от холодной тундры до жарких тропиков, отдельные виды освоили пресные и морские водоемы.
Покрытосеменные составляют большую часть массы растительного сообщества, являются звеном в цепи питания (продуцентами) – важнейшими производителями
органических веществ на суше, как водоросли – в морях и океанах.
В настоящее время цветковые господствуют на Земле. Такое доминирующее положение им позволили занять прогрессивные особенности:
- Возникновение цветка
- Защищенный семязачаток
- Двойное оплодотворение
- Возникновение плода
- Хорошо развита проводящая ткань
- Редукция гаметофита
- Опыление
- Многоярусные сообщества
Цветок – генеративный орган покрытосеменных (цветковых), высшая ступень полового размножения.
Цветок характерен только для покрытосеменных растений, ни один из других отделов подобным генеративным органом не обладает. По своему строению
цветок это видоизмененный обоеполый стробил, гомологичный стробилам голосеменных.
В отличие от голосеменных, у которых семязачатки лежат открыто на семенных чешуях, у цветковых семязачаток находится в замкнутом вместилище –
завязи, сформированной из плодолистика (-ов).
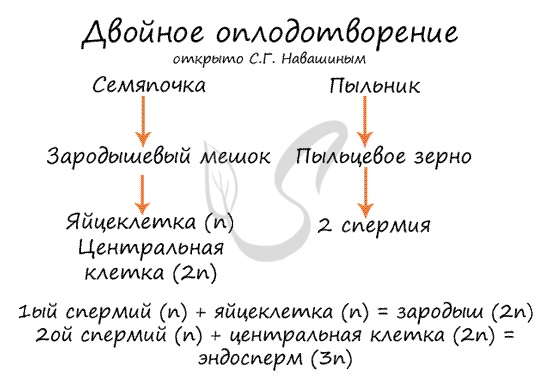
Двойное оплодотворение, открытое Навашиным Сергеем Гавриловичем, уникальное явление, характерное только для цветковых. Оно связано с тем, что в зародышевый мешок
попадают два спермия, один из которых (n) сливается с центральной клеткой (2n), с образованием запасного питательного вещества – эндосперма (3n).
Другой спермий (n) сливается с яйцеклеткой (n) с образованием зиготы (2n), из которой развивается зародыш.
У цветковых появляется плод – генеративный орган, служащий для защиты и распространения семян.
Ксилема – проводящая ткань, обеспечивающая восходящий ток воды и растворенных в ней минеральных солей, представлена не трахеидами,
а сосудами. Во флоэме ситовидные элементы окружены клетками-спутницами.
У покрытосеменных мы не найдем антеридиев и архегониев: гаметофиты максимально редуцированы.
В процессе опыления покрытосеменных участвуют насекомые, летучие мыши, птицы. Также опыление может происходить с помощью воды или ветра.

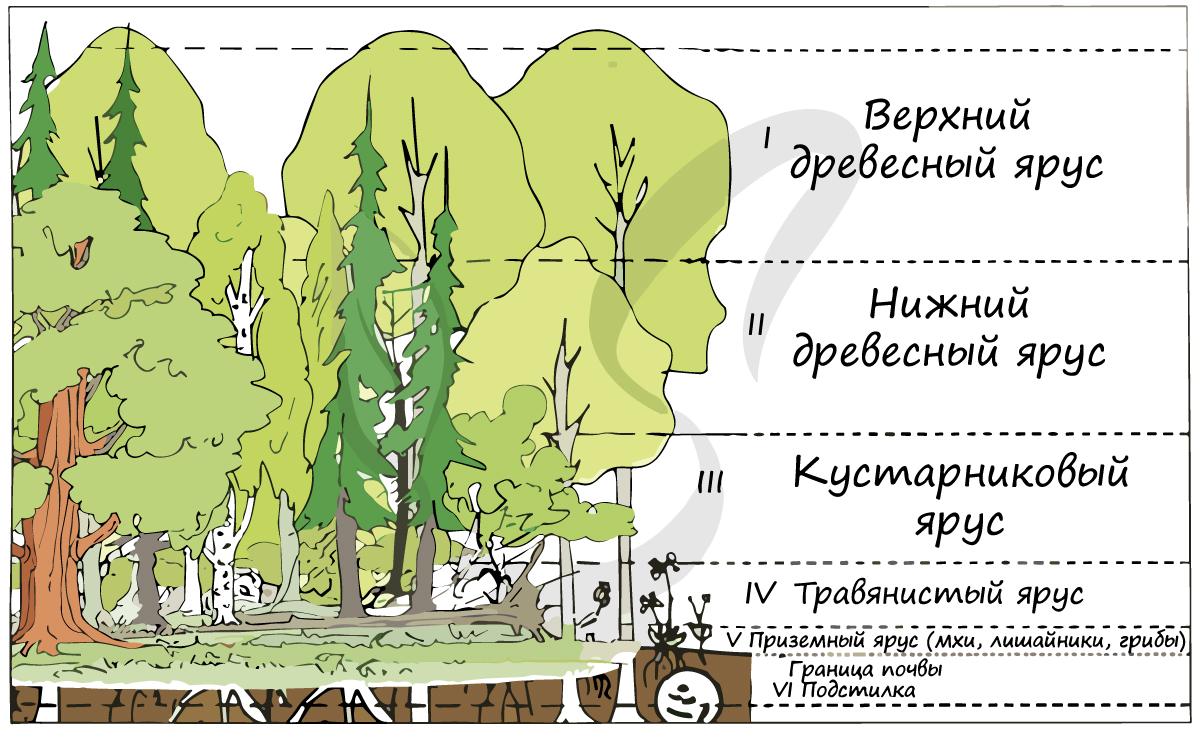
Особенностью цветковых является способность образовывать многоярусные сообщества, более устойчивые и продуктивные.
Многоярусность растительного сообщества служит приспособлением к равномерному распределению света: светолюбивые растения занимают верхний ярус, а теневыносливые растения отлично чувствуют себя в тени светолюбивых 🙂
Классы покрытосеменных
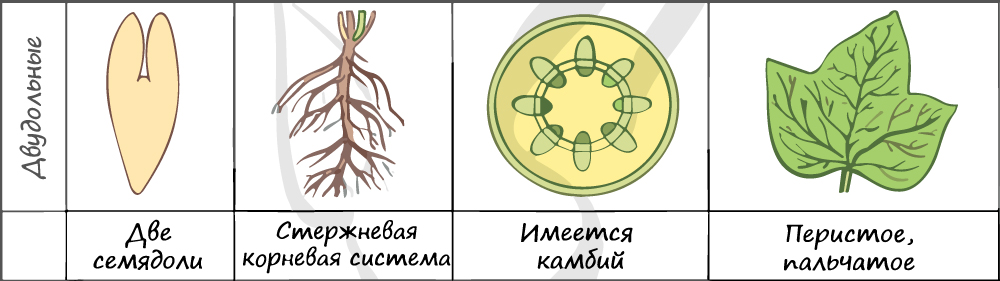
Отдел покрытосеменные состоит из двух классов: однодольные и двудольные. К классу двудольных относятся семейства: крестоцветные, сложноцветные, розоцветные,
бобовые (мотыльковые), пасленовые. Класс однодольные включает в себя семейства: злаковые, лилейные. Для каждого класса имеются характерные признаки.
- Двудольные – семейства: крестоцветные, сложноцветные, бобовые, розоцветные, пасленовые
- В составе зародыша обычно имеется две семядоли
- Листья
- Камбий
- Корневая система
- Цветок
- Однодольные – семейства: лилейные, злаки
- Зародыш содержит только одну семядолю
- Листья только простые, жилкование параллельное и дуговое
- Камбий отсутствует – из-за этого нет вторичного роста
- Корневая система мочковатая, главный корень и боковые рано отмирают, развиваются придаточные корни
- Цветок
- Жизненные формы – травы (древесные формы очень редки)
В семядолях содержится запас питательных веществ. При надземном прорастании семядоли (зародышевые листья) могут выполнять функцию фотосинтеза.

Листья двудольных простые и сложные, для двудольных характерно перистое и пальчатое жилкование.
За счет камбия растения растут в толщину, возможен вторичный рост осевых органов (стебля и корня).
Корневая система чаще всего стержневого типа, с хорошо выраженным главным корнем, от которого отходят боковые корни.
Главный корень развивается из зародышевого корешка.
Цветки пятичленные, реже встречаются четырехчленные. Хорошо обособлены чашечка и венчик.

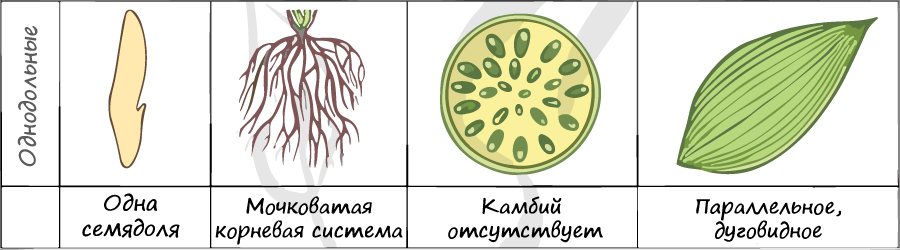
Цветок с простым околоцветником. Цветки чаще трехчленные, четырехчленные. Никогда не бывают пятичленными.
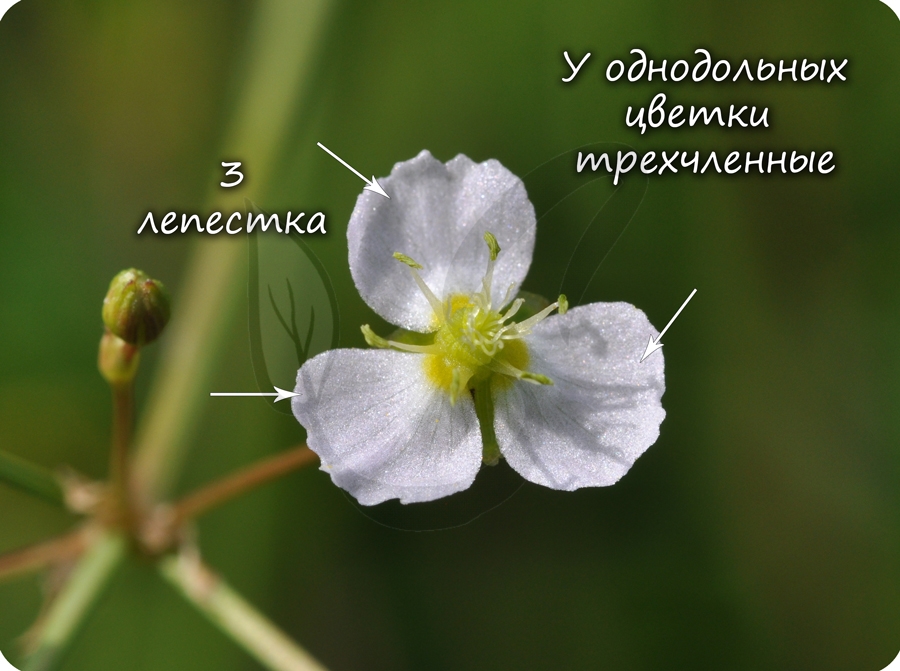
Эндосперм семени
Эндосперм (от греч. endon – внутри + греч. sperma – семя) – запасное питательное вещество, у покрытосеменных триплоидный (3n).
Эндосперм в семени есть у подавляющего большинства однодольных (лука, ландыша, пшеницы) и
двудольных (тмина, хурмы, фиалки). Отсутствует эндосперм в семенах тыквенных, крестоцветных (капусты), сложноцветных (подсолнечника), бобовых (гороха, фасоли), также у – березы, липы, дуба, клена, так как на ранней стадии развития растущий зародыш поглощает эндосперм.
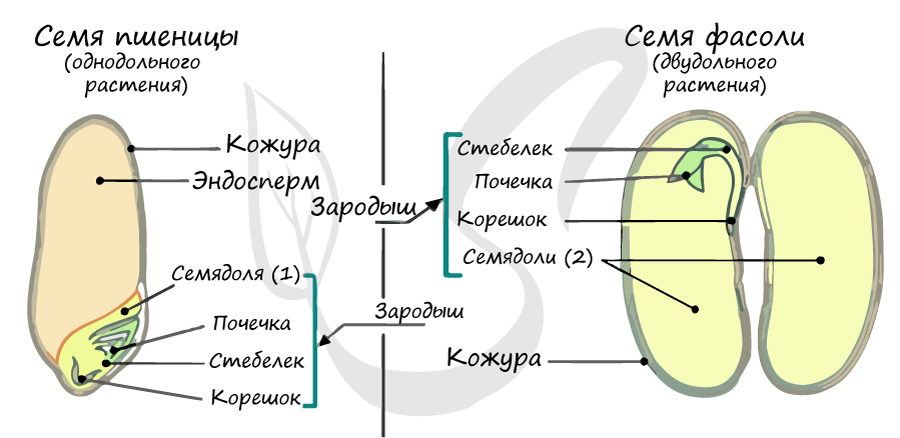
Жизненный цикл
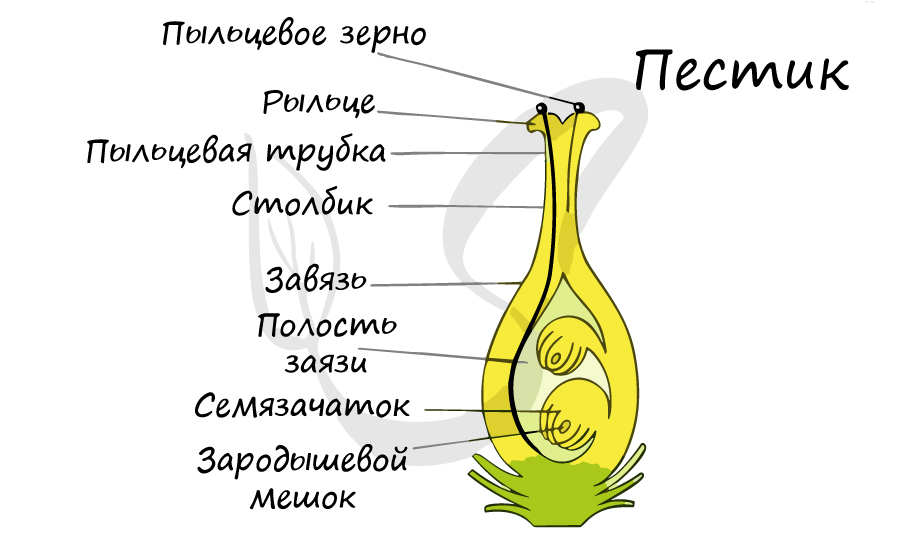
Из генеративных почек спорофита развиваются цветки. У взрослого растения спорофита (2n)
в цветке в гнездах пыльников тычинок в ходе микроспорогенеза образуется пыльцевое зерно (n) – мужской гаметофит. В завязи пестика в
семязачатке формируется женский
гаметофит – зародышевый мешок, внутри которого находятся центральная клетка (2n) и яйцеклетка (n).
В результате опыления (насекомым, ветром, человеком) пыльца с тычинок переносится на рыльце пестика. Пыльцевое зерно состоит из вегетативной и генеративной клеток.
Вегетативная клетка начинает растворять ткани пестика, образует пыльцевую трубку и прорастает до зародышевого мешка. Генеративная клетка делится, образуя
два спермия (n), из которых один сливается с центральной клеткой (2n) с образование эндосперма (3n) – запасного питательного вещества. Другой спермий (n)
сливается с яйцеклеткой (n), образуя зиготу (2n).
В дальнейшем из семязачатка формируется семя, а завязь превращается в околоплодник – образуется плод. Своим внешним видом плоды привлекают животных, и те их
охотно поедают) Благодаря семенной кожуре семена не подвергаются расщеплению в желудочно-кишечном тракте человека и животных. Они выходят из ЖКТ в неизменном виде и остаются способны к прорастанию: так происходит расселение растений. Попав в благоприятные условия,
они прорастают в спорофит (2n). Цикл замыкается.
Значение покрытосеменных
Покрытосеменным в жизни человека отведено важное место. Только подумайте – почти все культурные растения принадлежат к этому отделу! Цветковые имеют
медицинское значение, из многих растений изготавливаются лекарства. Их древесина используется для изготовления бумаги, мебели, применяются в промышленности.

© Беллевич Юрий Сергеевич 2018-2020
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Источник
Проводящие ткани служат для передвижения по растению растворенных в воде питательных — органических и неорганических — веществ. Подобно покровным тканям, они возникли как следствие приспособления растений к жизни в двух средах — почвенной и воздушной. В связи с этим появилась необходимость транспортировки питательных веществ в двух направлениях.Проводящие ткани служат для передвижения по растению растворенных в воде питательных — органических и неорганических — веществ. Подобно покровным тканям, они возникли как следствие приспособления растений к жизни в двух средах — почвенной и воздушной. В связи с этим появилась необходимость транспортировки питательных веществ в двух направлениях. От корня к листьям движется восходящий, или транспирационный, ток водных растворов солей. Ассимиляционный, нисходящий, ток органических веществ направляется от листьев к корням. Восходящий ток осуществляется почти исключительно по трахеальным элементам ксилемы, а нисходящий — по ситовидным элементам флоэмы. Сильно разветвленная сеть проводящих тканей несет водораство римые вещества и продукты фотосинтеза ко всем органам растения, начиная от тончайших корневых окончаний до самых молодых побегов. Проводящие ткани объединяют все органы растения в единую систему, Помимо дальнего, т. е. осевого, транспорта питательных веществ, по проводящим тканям частично осуществляется и ближний — радиальный транспорт. Все проводящие ткани являются сложными, или комплексными, т. е. состоят из морфологически и функционально разнородных элементов. Формируясь из одних и тех же меристем, два типа проводящих тканей — ксилема и флоэма — располагаются рядом. Во многих органах растений ксилема объединена с флоэмой в виде различного рода продольных тяжей, называемых проводящими пучками.Существуют первичные и вторичные проводящие ткани. Первичные ткани закладываются в листьях, молодых побегах и корнях. Они дифференцируются из клеток прокамбия. Вторичные проводящие ткани, обычно более мощные, возникают из камбия. {mospagebreak}Ксилема (древесина). Термин «ксилема» ввел немецкий ботаник К. В. Негели (1817-1891). По ксилеме от корня к листьям передвигаются вода и растворенные в ней минеральные вещества. Первичная и вторичная ксилемы содержат клетки одних и тех же типов. Однако первичная ксилема не формирует сердцевинных лучей, отличаясь этим от вторичной. Первичная ксилема формируется из прокамбия, вторичная — из камбия. В состав ксилемы входят морфологически различные элементы, осуществляющие функции как проведения, так и хранения запасных веществ, а также чисто опорные функции. Дальний транспорт осуществляется по трахеальным элементам ксилемы — трахеидам и сосудам, ближний в основном по паренхимным элементам. Дополнительные — опорную, а иногда и запасающую — функции выполняют трахеальные элементы и волокна механической ткани либрифор-ма, также входящие в состав ксилемы. Трахеиды в зрелом состоянии — это мертвые прозенхимные клетки, суженные на концах и лишенные протопласта. Длина трахеид в среднем составляет 1-4 мм, поперечник же не превышает десятых и даже сотых долей миллиметра. Стенки трахеид одревесневают, утолщаются и несут простые или окаймленные поры, через которые происходит фильтрация растворов, с помощью которой осуществляется дальний транспорт. Впрочем, боковые стенки трахеид в определенной степени водопроницаемы, что способствует осуществлению ближнего транспорта. Большая часть окаймленных пор находится около окончаний клеток, т. е. там, где растворы «просачиваются» из одной трахеиды в другую. Трахеиды есть у спорофитов всех растений, а у большинства хвощевидных, плауновидных, папоротнико-видных и голосемянных они являются единственными проводящими элементами ксилемы. Между трахе-идами и волокнами либриформа существуют переходные формы. Сосуды — это полые трубки, состоящие из отдельных члеников, располагающихся друг над другом. Между расположенными один над другим члениками одного и того же сосуда имеются разного типа сквозные отверстия — перфорации. Благодаря перфорациям вдоль всего сосуда свободно осуществляется ток жидкости. Эволюционно сосуды, по-видимому, произошли из трахеид путем разрушения замыкающих пле нок пор и последующего их слияния в одну или несколько перфораций. Концы трахеид, первоначально сильно скошенные, заняли горизонтальное положение, а сами трахеиды стали короче и превратились в членики сосудов. Сосуды могут состоять из весьма значительного числа члеников различной длины и диаметра. Общая же длина сосудов достигает иногда нескольких метров. Диаметр же варьирует от 0,2 мм до 1 мм. Последнее зависит от вида растения, а у деревьев, растущих в сезонном климате, также и от того, в какой части ксилемы — «весенней» или «осенней» — сосуд расположен. Сосуды появились независимо в разных линиях эволюции наземных растений. Однако наибольшего развития они достигают у покрытосемянных, где являются главнейшими водопроводящими элементами ксилемы. Возникновение сосудов — важное свидетельство эволюционного прогресса этого таксона, поскольку они существенно облегчают транспирационный ток вдоль тела растения. Помимо первичной оболочки, сосуды, как и многие трахеиды, в большинстве случаев имеют вторичные утолщения. В самых ранних трахе-альных элементах вторичная оболочка может иметь форму колец, не связанных друг с другом {кольчатые трахеиды и сосуды). Позднее появляются трахеальные элементы со спиральными утолщениями. Затем следуют сосуды и трахеиды с утолщениями, которые могут быть охарактеризованы как спирали, витки которых связаны между собой {лестничные утолщения). В конечном итоге вторичная оболочка сливается в более или менее сплошной цилиндр, формирующийся внутрь от первичной оболочки. Этот цилиндр прерывается в отдельных участках порами. Сосуды и трахеиды с относительно небольшими округлыми участками первичной клеточной оболочки, не прикрытыми изнутри вторичной оболочкой, нередко называют пористыми. В тех случаях, когда поры во вторичной оболочке образуют подобие сетки или лестницы, говорят о сетчатых или лестничных трахеальных элементах (лестничные сосуды и трахеиды). Вторичная, а иногда и первичная оболочки, как правило, лигнифици-руются, т. е. пропитываются лигнином, это придает дополнительную прочность, но ограничивает возможности дальнейшего их роста в длину. Трахеальные элементы, т. е. трахеиды и сосуды, распределяются в ксилеме различным образом. Иногда на поперечном срезе они образуют хорошо выраженные кольца {кольце-сосудистая древесина}. В других случаях сосуды рассеяны более или менее равномерно по всей массе ксилемы (рассеяннососудистая древесина}. Особенности распределения трахеальных элементов в ксилеме используют при определении древесины различных пород деревьев. Помимо трахеальных элементов, ксилема включает лучевые элементы, т. е. клетки, образующие сердцевинные лучи, сформированные чаще всего тонкостенными клетками (лучевая паренхима). Реже, например, в лучах хвойных, встречаются лучевые трахеиды. По сердцевинным лучам осуществляется ближний транспорт веществ в горизонтальном направлении. В ксилеме покрытосемянных, помимо проводящих элементов, содержатся также тонкостенные неодревесневшие живые паренхимные клетки, называемые древесинной паренхимой. По ним отчасти наряду с сердцевинными лучами осуществляется ближний транспорт. Кроме того, древесинная паренхима служит местом хранения запасных веществ. Элементы сердцевинных лучей и древесинной паренхимы, подобно трахеаль-ным элементам, возникают из камбия, но из паренхимных инициалей. Клетки паренхимы, примыкающие к сосуду, могут (обычно у деревьев) образовывать выросты в полость сосуда через поры, так называемые тилы. Иногда тилы заполняют всю полость сосуда, и в этом случае проводящая функция нарушается. Тилообразование усиливает механическую прочность центральной части стволов деревьев. Кроме того, тилы играют особую роль в процессе формирования ядра древесины. {mospagebreak}Флоэма. Термин «флоэма» ввел К. В. Негели в 1858 г. Флоэма — сложная проводящая ткань, по которой осуществляется транспорт продуктов фотосинтеза от листьев к местам их использования или отложения (к точкам роста, подземным органам, зреющим семенам и плодам и т. д.). Первичная флоэма дифференцируется из прокамбия, вторичная (луб) — производное камбия. В стеб лях флоэма располагается обычно снаружи от ксилемы, а в листьях обращена к нижней стороне пластинки. Первичная и вторичная флоэмы, помимо различной мощности ситовидных элементов, отличаются тем, что у первой отсутствуют сердцевинные лучи. В состав флоэмы входят ситовидные элементы, паренхимные клетки, элементы сердцевинных лучей и механические элементы. Большинство клеток нормально функционирующей флоэмы живые. Отмирает лишь часть механических элементов. Собственно проводящую функцию осуществляют ситовидные элементы. Различают два их типа: ситовидные клетки и ситовидные трубки. Стенки ситовидных элементов содержат многочисленные мелкие сквозные канальцы, собранные группами в так называемые ситовидные поля. У ситовидных клеток, вытянутых в длину и имеющихзаостренные концы, ситовидные поля располагаются главным образом на боковых стенках. Ситовидные клетки — основной проводящий элемент флоэмы у всех групп растений, исключая покрытосемянные. Клеток-спутниц у ситовидных клеток нет. Ситовидные трубки покрытосемянных более совершенны. Они состоят из отдельных клеток — члеников, располагающихся один над другим. Длина отдельных члеников ситовидных трубок колеблется в пределах 150-300 мкм. Поперечник ситовидных трубок составляет 20-30 мкм. Эволюционно их членики возникли из ситовидных клеток. Ситовидные поля этих члеников находятся главным образом на их концах. Ситовидные поля двух расположенных один над другим члеников образуют ситовидную пластинку. Ситовидные поля (название указывает на их сходство с ситом) представляют собой участки клеточной стен ки, пронизанные многочисленными мелкими отверстиями, через которые с помощью цитоплазматических тяжей сообщаются протопласты соседних ситовидных элементов. Членики ситовидных трубок формируются из вытянутых клеток прокамбия или камбия. При этом материнская клетка меристемы делится в продольном направлении и производит две клетки. Одна из них превращается в членик, другая — в клетку-спутницу. Наблюдается и поперечное деление клетки-спутницы с последующим образованием двух-трех подобных клеток, расположенных продольно одна над другой рядом с члеником. Предполагается, что клетки-спутницы вместе с члениками ситовидных трубок составляют единую физиологическую систему и способствуют продвижению тока асси-милятов. Кроме того, в клетках-спутницах вырабатываются различные ферменты, которые передаются в ситовидные трубки. При своем формировании членик ситовидной трубки имеет постенную цитоплазму, ядро и вакуоль. С началом функциональной деятельности он заметно вытягивается. На поперечных стенках появляется множество мелких отверстий-перфораций, образующих канальцы диаметром в несколько микрометров, через которые из членика в членик проходят цитоплазматиче-ские тяжи. На стенках канальцев откладывается полисахарид — каллоза, сужающий их просвет, но не прерывающий цитоплазматическиетяжи. По мере развития членика ситовидной трубки в протопласте образуются слизевые тельца. Ядро и лейкопласты, как правило, растворяются. Граница между цитоплазмой и вакуолью — тонопласт — исчезает, и все живое содержимое сливается в единую массу. При этом цитоплазма теряет полупроницаемость и становится вполне проницаемой для растворов органических и неорганических веществ. Слизевые тельца также теряют очертания, сливаются, образуя слизевый тяж и скопления около ситовидных пластинок. На этом формирование членика ситовидной трубки завершается. Длительность функционирования ситовидных трубок невелика. У кустарников и деревьев она продолжается не более 3-4 лет. По мере старения ситовидные трубки закупориваются каллозой (образующей так называемое мозолистое тело} и затем отмирают. Отмершие ситовидные трубки обычно сплющиваются давящими на них соседними живыми клетками. Основными веществами флоэмного тока являются сахара, главным образом сахароза. Кроме того, обнаружены азотсодержащие вещества (преимущественно аминокислоты), органические кислоты и фито-гормоны. Скорость передвижения ассими-лятов во флоэме относительно невелика — 50-100 см/ч (по ксилеме примерно в 2 раза быстрее). Сам процесс передвижения включает ряд этапов и идет со значительной затратой энергии. Паренхимные элементы флоэмы [лубяная паренхима} состоят из тонкостенных клеток. В них откладываются запасные питательные вещества, и отчасти по ним осуществляется ближний транспорт ассимиля-тов. У голосемянных клетки-спутницы отсутствуют, и их роль до известной степени выполняют прилегающие к ситовидным клеткам немногочисленные клетки лубяной паренхимы. Сердцевинные лучи флоэмы также состоят из тонкостенных парен-химных клеток. Они предназначены для осуществления ближнего транспорта ассимилятов. {mospagebreak}Проводящие пучки. Обособленные тяжи проводящей системы, состоящие чаще из ксилемы и флоэмы, называют проводящими пучка-ми. Первоначально они возникают из прокамбия. Из клеток прокамбия вначале дифференцируются элементы протофлоэмы (центробежно) и протоксилемы (центростремительно). У корня и те и другие элементы дифференцируются центростремительно. Позднее прокамбий образует элементы метафлоэмы и метакси-лемы. Образовавшиеся из прокамбия проводящие пучки иногда называют первичными. В тех случаях, когда часть прокамбия сохраняется и превращается затем в камбий, а пучок способен к вторичному утолщению, говорят об открытых пучках. Они встречаются у большинства двудольных и голосемянных. В закрытых пучках однодольных прокамбий полностью дифференцируется в проводящие ткани и далее не утолщается. Вокруг пучков нередко формируется обкладка из живых или мертвых паренхимных клеток. Они могут быть полными, т. е. состоящими из флоэмы и ксилемы, или изредка неполными, состоящими только из ксилемы или флоэмы. В зависимости от взаимного расположения флоэмы и ксилемы различают пучки нескольких типов. Чаще всего флоэма лежит по одну сторону от ксилемы. Такие пучки называют коллатеральными (открытые и закрытые). У части двудольных растений (из семейств пасленовых, вьюнковых, тыквенных и т. д.) одна, более мощная, часть флоэмы располагается снаружи от ксилемы (камбий располагается между ними), а другая — с внутренней стороны ксилемы. Такой пучок называется биколлатеральным, а соответствующие участки флоэмы — наружной и внутренней флоэмой. Биколла-теральные пучки формируются, оче- • видно, в результате слияния двух коллатеральных пучков. Встречаются также концентрические пучки, при этом флоэма окружает ксилему (центроксилемные пучки) либо, наоборот, ксилема окружает флоэму (центрофлоэмные). Центрофлоэмные пучки найдены в стеблях и корневищах ряда двудольных (ревень, щавель,бегония)и однодольных (многие лилейные, осоковые). Известны пучки промежуточные между закрытыми коллатеральными и центрофлоэмными. Центроксилемные пучки обычны для папоротников. В центре молодых корней голосемянных и покрытосемянных, имеющих первичное строение, располагается проводящий пучок, получивший название радиального. Ксилема в таком пучке как бы расходится лучами от центра, а флоэма располагается между лучами. Возникают эти пучки из прокамбия. Встречаются однолучевые (монархные), двулучевые (диархные), трехлучевые (триар-хные), четырехлучевые (тетрархные), пятилучевые (пентархные) и многолучевые (полиархные) радиальные пучки. Последние обычны у однодольных.
29.06.2015
Источник