
Продукты обмена у насекомых выделяются через мальпигиевы сосуды

Выделительная система – система органов, отвечающая за выведение из организма конечных продуктов обмена веществ.

Выделительная система
Выделительная система

1 – средняя кишка, 2 – мальпигиевы сосуды,
3 – задняя кишка, 4 – прямая кишка
Использовано изображение:[4]
Мальпигиевы сосуды: строение и функция
Главными органами выделения являются мальпигиевы сосуды. Так называются тонкие длинные трубочки, которые впадают в кишечный канал в области соединения средней и задней кишок. Их количество составляет от 1 до 250 и даже более. Если их много, они обычно группируются в пучки.[2]
Основное количество метаболитов, выводимых этими органами, образуются в процессе обмена белков; это различные производные аминокислот, мочевина, мочевая кислота, аммиак. Они синтезируются в клетках и выбрасываются в гемолимфу, откуда, в свою очередь, извлекаются мальпигиевыми сосудами. Последние открываются и выводят свое содержимое в кишечный канал. Отведение осуществляется благодаря тому, что в стенках мальпигиевых сосудов имеются мышечные пучки, вызывающие их сокращение.[3](фото)
У примитивных насекомых (коллемболы, тли, некоторые двухвостки) этих органов выделения нет, а у трипсов, уховерток и щетинохвосток в них нет мышечного слоя. Проникая в гемолимфу, сосуды лишь пассивно колеблются в ее токах, и так же пассивно и неполноценно (в сравнении с высшими насекомыми) выполняют свою функцию. У прямокрылых мальпигиевы сосуды также устроены довольно просто. Они насасывают внутрь себя плазму, не отделяя от нее вредных веществ. Потом эта «первичная моча» выбрасывается в кишечник, и уже там жидкая часть плазмы и соединения, необходимые организму, всасываются обратно, а метаболиты, которые надо вывести, остаются. Плохая «производительность» таких сосудов компенсируется у этих насекомых их большим числом.В более сложных случаях в сосуды всасываются только вещества, подлежащие выведению.[1]

Личинка муравьиного льва
Личинка муравьиного льва
Использовано изображение:[5]
Особенности работы выделительной системы
У многих насекомых мальпигиевы сосуды «работают» постоянно, но в ряде случаев их функция может периодически усиливаться под действием определенных стимулирующих факторов.[1]
Например, у клопа Rhodniusprolixus, который за один раз насасывает большое количество крови и после питания нуждается в интенсивном выведении метаболитов, имеется гормональный механизм, регулирующий работу органов выделения. Грудные ганглии этих насекомых выделяют диуретический гормон, под действием которого скорость экскреции продуктов обмена увеличивается в 1000 раз. Образование этого гормона происходит в тот момент, когда брюшко клопа начинает растягиваться во время питания.[1]
Иногда мальпигиевы сосуды берут на себя не только выведение экскретов, но и добавочные функции. Например, у личинок муравьиного льва (фото) перед окукливанием они превращаются в шелкоотделительные железы. Образующийся шелк скапливается в прямой кишке, а затем постепенно выходит наружу, и насекомое плетет из него свой кокон.[3]

Гусеница Hyalophora cecropia
Гусеница Hyalophora cecropia

Использовано изображение:[6]
Другие органы, участвующие в выведении продуктов метаболизма
Летучие и нелетучие продукты обмена веществ, а также балластные и отравляющие вещества, проникшие в организм насекомого извне, удаляются из него сразу несколькими путями. Несмотря на то, что не все из перечисленных ниже органов относятся собственно к выделительной системе, все они участвуют в «очищении» тела насекомого. Итак, в широком понимании к органам выделения можно отнести:
Мальпигиевы сосуды (своеобразный аналог почек животных) – выводят метаболиты из тканей;
Кишечник – освобождает организм от непереваренных остатков пищи и некоторых продуктов обмена (мочевая кислота, мочевина);
Трахейная система – удаляет из организма углекислоту;[1][3]
Железы. Например, у коллебмол функцию экскреции выполняют лабиальные железы, а у гусеницы шелкопряда Hyalophora cecropia (фото) выводить продукты обмена могут шелкоотделительные железы. У самцов ряда тараканов мочевая кислота выделяется через половые железы, которыми она используется для покрытия сперматофоров.[3]
Также в выведении летучих веществ участвуют покровы тела, а некоторые ядовитые соединения накапливаются в жировом теле и кутикуле, что также делает их участниками процесса выведения из организма всего «лишнего».[1][3] Так, у американской белой бабочки кристаллы мочевой кислоты накапливаются в чешуйках крыльев, из-за чего ее крылья имеют характерный белоснежный цвет.[3]
Статья составлена с использованием следующих материалов:
Литературные источники:
1.
Захваткин Ю.А., Курс общей энтомологии, Москва, «Колос», 2001 – 376 с.
2.
Росс Г., Росс Ч., Росс Д. Энтомология. – М., Мир, 1985. -572 с.
3.
Шванвич Б.Н. Курс общей энтомологии. – М.Л. Советская наука. 1949.-900 с., ил.
Изображения (переработаны):
4.
Догель В.А. Зоология беспозвоночных. /Под ред. проф. Полянского Ю. И. – 7-е изд., перераб. и доп. – М.: Высш.школа., 1981. – 606 с., Иллюстрации из книги ©
5.
6.
Свернуть Список всех источников
Источник
Выделительная системаВыделительная система насекомых сложна, многообразна и может быть подразделена на три отдельные группы органов или желез – экскреторную систему, секреторную систему и эндокринную систему. Общим их физиологическим свойством является способность выделять наружу или внутрь разнообразные вещества и тем самым участвовать в обмене веществ в организме. В одних случаях образуются вредные или бесполезные продукты обмена веществ, и они удаляются из организма, в других случаях разнообразные органы и железы выделяют необходимые для нормальной жизнедеятельности вещества. Экскреторная система и экскреция. В процессе переваривания пищи и использования усвоенных веществ в организме образуются ненужные или вредные для него вещества. Удалением непереваренных и неиспользованных остатков пищи организм лишь частично освобождается от этих веществ. Большое количество бесполезных или вредных веществ образуется в результате жизненных процессов в тканях и органах, т. е. вне пищеварительной системы. Экскреторная, или собственно выделительная, система и обеспечивает удаление из организма таких веществ; тем самым она поддерживает относительное биохимическое постоянство его внутренней среды и контролирует ионный состав крови. Эти ненужные или вредные вещества, удаляемые из организма, называются экскретами, а сам процесс их выделения – экскрецией. Основными выделяемыми веществами являются азотистые вещества; они возникают в организме в результате усвоения и распада белков пищи. Действительно, при окислении безазотистых компонентов пищи – углеводов и жиров – конечными продуктами служат углекислый газ (СО2) и вода, которые удаляются из организма с помощью дыхательной системы. Совсем другое положение создается при окислении белков; помимо СО2 и воды, образуются вредные азотсодержащие вещества, накопление которых в организме могло бы повести к его гибели. Отсюда и возникает необходимость в выделительной системе, освобождающей организм от таких азотсодержащих веществ, а также от катионов калия, натрия, кальция, магния и от кислот щавелевой, фосфорной и др. Главнейшим органом выделения служат мальпигиевы сосуды: названы они так по имени итальянского ученого Мальпиги, впервые открывшего их у насекомых еще в XVII в. Это слепые на свободном конце трубочки, прикрепленные своим основанием к кишечнику на границе между средней кишкой и задней (рис. 22). Стенки мальпигиевых сосудов состоят изнутри из одного слоя эпителиальных клеток, снаружи одеты базальной перепонкой (рис. 29) и нередко имеют собственную мускулатуру, обеспечивающую им подвижность. Число мальпигиевых сосудов у разных насекомых колеблется в пределах 2-200 и более или менее характерно для разных систематических групп. Такие низшие группы крылатых насекомых, как стрекозы и прямокрылые, имеют от 30-50 до 200 трубочек, тогда как клопы, вши, двукрылые и блохи имеют всего 4 трубочки, жуки – 4-6, ручейники и бабочки – 6. Однако некоторые специализованные низшие группы имеют уменьшенное число трубочек, например 2-8 у термитов и 8-20 у уховерток; с другой стороны, у многих жалящих перепончатокрылых их насчитывается более 100. Наконец, в немногих случаях, как у тлей и некоторых первичнобескрылых, мальпигиевы сосуды отсутствуют.
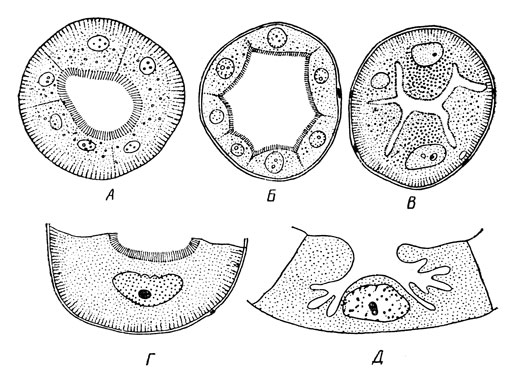
Рис. 29. Поперечный разрез через отдельные мальпигиевы сосуды: А – медведки; Б – медоносной пчелы; В – плавунца, диетальная часть; Г и Д – личинки обыкновенного муравьиного льва, отдельные клетки (Г – из основания, Д – из вершинной части) (из Н. Я. Кузнецова) Своей свободной частью мальпигиевы сосуды взвешены и как бы плавают в гемолимфе, отсасывая из нее продукты выделения. Однако у гусениц бабочек, у многих жуков и их личинок и у некоторых других насекомых конец трубочек фиксирован на задней кишке; это явление обозначается как криптонефрия и обеспечивает всасывание трубочками воды из задней кишки, тем самым дополняя функцию последней. Омывая мальпигиевы сосуды, гемолимфа отдает им накапливающиеся в ней продукты обмена: азотистые вещества преимущественно в виде мочевой кислоты и ее солей – уратов, а также различные неорганические ионы. Вещества эти диффундируют в мальпигиевы сосуды, откуда выводятся в кишечник и удаляются вместе с экскрементами через анальное отверстие. В общем мальпигиевы сосуды имеют выносящую экскреторную функцию и аналогичны почкам позвоночных животных. Однако в некоторых случаях мальпигиевы сосуды имеют дополнительную – уже секреторную – функцию, выделяя необходимые организму вещества. Особенно замечательно выделение прядильных веществ (шелка и иногда примесей к нему), идущих на образование кокона перед окуклением у личинок настоящих сетчатокрылых и некоторых жуков. Следовательно, эта функция мальпигиевых сосудов аналогична функции шелкоотделительных (слюнных) желез у гусениц бабочек. Известны также случаи выделения у отдельных видов насекомых некоторых пищеварительных ферментов и других веществ. Выносящую экскреторную функцию имеют также нижнегубные, или лабиальные, железы, свойственные первичнобескрылым насекомым – подурам, двухвосткам и щетинохвосткам. Это парные образования с общим каналом, открывающимся у основания нижней губы. Они интересны филогенетически, так как имеются и у ракообразных и сравниваются с метанефридиями кольчатых червей. Следует отметить, что у подур и некоторых двухвосток мальпигиевы сосуды полностью отсутствуют. Экскреторную функцию имеет и жировое тело, хотя важнейшая его роль состоит в накоплении в организме запасных питательных веществ. Но в отличие от выносящей функции мальпигиевых сосудов и лабиальных органов, жировое тело производит внутриклеточное накопление экскретов, обычно в виде кристаллов. Эти экскреты остаются в жировом теле либо пожизненно, либо после накопления передаются мальпигиевым сосудам, которые и выводят их из организма. Последний способ свойствен личинкам стебельчатых перепончатокрылых и высших двукрылых, а также гусеницам бабочек; освобождение жирового тела от экскретов происходит у них при окуклении. Экскреторную накопляющую функцию приписывают также нефроцитам – группам клеток, способным поглощать из полости тела введенные туда посторонние вещества: аммиачный кармин, белки, хлорофилл и другие вещества. Наиболее заметная совокупность нефроцитов локализована около спинного сосуда в перикардиальной полости; отсюда они и называются перикардиальными клетками (рис. 20). Функцию нефроцитов исследовал в конце XIX в. А. О. Ковалевский, но в настоящее время их значение в процессе выделения остается неясным, так как в течение жизни насекомого в них не происходит накопления каких-либо веществ. Выделительным процессом можно считать также и линьку, при которой организм освобождается от азотистых веществ; в этом случае хитин и основные красящие вещества – меланины – должны рассматриваться как экскреты. Экзокринные железы и секреция. Разнообразные железы насекомых выделяют вещества, так или иначе используемые организмом. Такие вещества обозначаются секретами, а сам процесс их выделения называется секрецией. Различают два основных типа секреции: выделение с помощью типичных желез, снабженных выводными протоками, когда секреты поступают в различные органы или полости либо наружу, и выделение непосредственно в кровь особыми железами, лишенными выводных протоков. В первом случае имеет место обычная секреция, и соответствующие железы обозначают понятием экзокринных желез; другой тип желез получил название эндокринных, а выделяемые ими вещества – гормонов. Оба эти способа выделения по своему физиологическому значению оказались столь различными, что их необходимо рассматривать отдельно. Экзокринные железы разнообразны по функциям и строению и частью уже рассматривались выше. Это различные железы как кожного (эктодермального), так и другого происхождения. Одни из них участвуют в пищеварении (слюнные и железы средней кишки), другие выделяют вещества механической защиты (восковые, лаковые, шелкоотделительные), третьи образуют биологически активные вещества, обеспечивающие химическую защиту насекомого, четвертые выделяют привлекающие вещества. Примером желез, выполняющих защитную функцию, может быть ядовитая железа самок жалящих перепончатокрылых – пчел, ос, муравьев; ее выводной проток связан с жалом, представляющим собой видоизмененный яйцеклад. Ядовитый секрет перепончатокрылых обнаруживает свойства белковых ядов, вызывает сильное болевое чувство и при одновременном многократном ужалении может вызвать лихорадочное состояние с повышением температуры тела. В последнее время пчелиный яд успешно применяется в лечебных целях против ревматизма и сходных заболеваний. Есть также отпугивающие железы, выделяющие сильно пахнущие и другие отталкивающие вещества – репелленты. У жуков-бомбандиров (Brachinus, сем. жужелиц) есть пара желез, открывающихся у анального отверстия. При опасности у этих жуков выделяется из желез секрет, содержащий азотнокислые соли и окись азота; в смеси с воздухом этот секрет взрывается с образованием маленького облачка. Отпугивающее значение приписывают также пахучим железам клопов, располагающихся на груди или брюшке; однако Д. М. Федотов показал, что у клопа-черепашки (Eurygaster integriceps Put.) эти железы выполняют в период миграции клопов привлекающую функцию, обеспечивая встречу полов и скопление клопов. В этом случае выделяемый пахучий секрет уже играет роль привлекающего вещества, или аттрактанта. Привлекающие железы, выполняющие сходную функцию, есть также и у ряда других насекомых – у самцов некоторых кузнечиков, сверчков и тараканов, у самок или самцов бабочек и др. В этих случаях выделяемое пахучее вещество играет роль полового аттрактанта, облегчающего встречу самца и самки одного вида. В целом с помощью аттрактантов обеспечивается распознавание особей своего вида, концентрация их на подходящих местах и заселение территории, встреча полов и пр. Следовательно, аттрактанты являются веществами для передачи информации, т. е. играют роль своеобразного химического языка, который обеспечивает связь между особями своего вида. Особую роль играют химические стимуляторы, выделяемые насекомыми – феромоны, или экзогормоны. Они воздействуют на поведение и развитие особей, по физиологическому эффекту сходны с гормонами, но не имеют эндокринного происхождения. Действием феромонов объясняют поведение рабочих особей у таких общественных насекомых, как муравьи и термиты и пр., т. е. эти вещества также участвуют в передаче химической информации. Эндокринные железы и внутренняя секреция. Как уже указывалось, эндокринные железы лишены выводных протоков и выделяют свою секреторную продукцию в кровь. Выделяемые вещества здесь называются гормонами, или инкретами, а сама секреция обозначается как внутренняя секреция, или инкреция. Попадая в кровь, гормоны транспортируются ею во все части тела. В целом эндокринные железы регулируют развитие насекомых и связанные с ними явления – личиночный рост, линьки, торможение развития (диапауза), половое созревание и пр. Наиболее изучены следующие четыре вида эндокринных желез: нейросекреторные клетки головного мозга, кардиальные тела, прилежащие тела и переднегрудные железы (рис. 30).
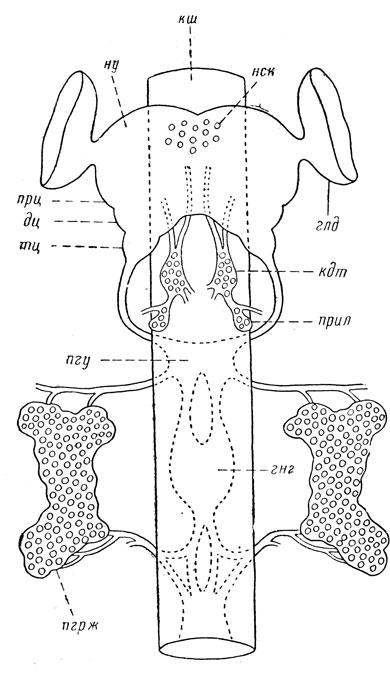
Рис. 30. Схема эндокринной системы насекомых и переднего отдела центральной нервной системы (ориг.): кш – кишечник, ну – надглоточный узел, нск – нейросекреторные клетки головного мозга, прц – протоцеребрум, дц – дейтоцеребрум, тц – тритоцеребрум, глд – глазные доли, пгу – подглоточный узел, кдт – кардиальные тела, прил – прилежащие тела, пгрж – переднегрудная железа, гнг – ганглии брюшной нервной цепочки Нейросекреторные клетки головного мозга выделяют мозговой гормон, функция которого, видимо, разнообразна. В ряде случаев этот гормон контролирует и активизирует деятельность переднегрудных желез; при прекращении его выделения у личинок и куколок затормаживается рост и развитие, возникает диапауза. Активационная роль мозгового гормона особенно хорошо изучена у шелкопряда Platysamia и у других чешуекрылых. Мозговой гормон может аккумулироваться в прилежащих и кардиальных телах, также способствуя их активизации. Переднегрудные, или проторакальные, железы представляют собой пару желез, расположенных в брюшной части переднегруди по бокам переднегрудного ганглия и связанных с последним нервами. Эти железы свойственны личинкам многих насекомых и выделяют линочный гормон, или экдизон; его эмпирическая формула С18Н30О4,и он был выделен химически в кристаллической форме. Экдизон прекращает диапаузу, вызывает у личинки линьку и регулирует, следовательно, развитие личинки. Прилежащие, или добавочные, тела (corpora allata) располагаются над передней кишкой позади головного мозга, обычно в виде пары округлых образований. Они выделяют ювенильный гормон, или неотенин, свойственны всем крылатым насекомым и части двухвосток (Diplura) как в личиночном, так и во взрослом состоянии. Действие ювенильного гормона состоит в том, что он способствует развитию личиночных органов и препятствует превращению во взрослую фазу, т. е. является ингибитором метаморфоза. При пересадке прилежащих тел в личинку последнего возраста линька во взрослую фазу замещается обычной линькой, и вместо взрослого насекомого возникает ненормально крупная личинка. Изменение действия ювенильного гормона в течение развития личинки объясняют падением его концентрации в крови к последнему возрасту личинки, вследствие чего роль его затухает. Происходит это потому, что рост тела личинки опережает рост прилежащих тел, развитие личиночных органов ослабевает и затем останавливается. В связи с этим происходит ослабление роли проторакальных желез как личиночного органа; прекращается выделение линочного гормона, линьки прекращаются, что и составляет характерную особенность взрослого насекомого. Минимальная концентрация ювенильного гормона отмечена у диапаузирующих куколок. Кардиальные тела (corpora cardiaca) сходны по форме и положению с прилежащими телами, близко примыкают к ним, но расположены впереди последних, связаны нервами с головным мозгом и свойственны личинкам и имаго. Их роль наименее изучена; у тараканов они регулируют секреторную деятельность проторакальных желез. Личинки двукрылых (Diptera) имеют иные особенности эндокринной системы. Она у них представлена так называемой кольцевой железой, расположенной позади мозга в виде небольшого кольца вокруг аорты. Верхняя часть этой железы имеет железистые клетки, соответствующие прилежащим телам, а нижняя – кардиальным телам (рис. 31).
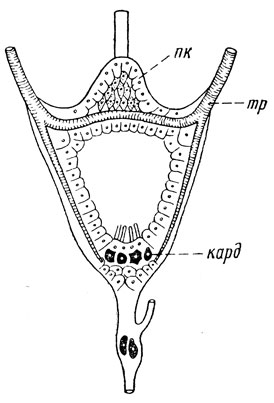
Рис. 31. Кольцевая железа личинки мухи Calliphora (по Томсену из Иммса): пк – прилежащие клетки, тр – трахея, кард – кардиальные клетки Эндокринные железы связаны друг с другом, действуют взаимосвязано, но функционируют также под воздействием нервной системы и внешней среды (тепло, свет и другие факторы). В целом эти железы образуют эндокринную систему, играющую важную роль в регуляции роста и развития насекомых. |
Источник