
Рецепторы внутренних органов сосудов
– . – .
, , .. .. . , .
, , . , – . , , , . , .
, . , ,
, , ( ), , , : , , , . , – , .
, . , , , . .
, , , , , ( ).
, ( ) , – – , . , , , – . , , ” “. , () : , , , . – , , – .
– ( , , ), , . , . ; , , . , . , , .-. , – , , , – . . . . , , , , . . , , – – , . , , , , , , . , – . , : – , , . , ( ) . , .
. , , ( ) , . .
“” , , , ( , ). : , , . , , ( ). .
..
. , , , . , , . , , .
. , () . , ( ), .
. , . , . . . , , . , , . : , , , . , . , .
: 1) , , . 2) , , . 3) – , , , . , . : ; . , , .
, . , . , , – .
. , , . , , . () : , . , . () , , , . () . , , . ( – – , ), .
, , . , . , , , .
( ) , .
.
. : (prosencephalon), (mesencephalon) (metencephalon). (telencephalon) (diencephalon). . (metencephalon) (myelencephalon), .
, – , , , – , , , , (, ). – , , ( , ). , , , , , , , ( ), ( , ). .
, , . , . ( ) . (, , ) , . , .
, , , . – – . , , , . , , , (, , , ), ( ). . , . , – . , , Th4 4 , 4 5 , 4 , , L5 , ( – L5 – S1, ), , , , . , , Th4 4 (I, XIV, III, IV, VII, VIII, XI), L5 (III, VII, IX).
…
. . , .
, . – – , . . , ( ). , .
– , – . , , . , , . , . , ( , . ), , . , ( ).
, , , , .
– – . – , , . , .
“” (.. , 1948), .
.
. , . . “”. “” , , (.. , 1965). – , , – , . , – , , – , .
. . , . , , . , ( – ). , .
.
…
() . (medulla spinalis) 41 – 45 , . , . , , . , .
…
. , . , . . , () , (, 1953, , 1956, , 1957). , ( ), , .
…
, , , . , , . , . , , . .
, , . 8, 12, 5, 5, 2 3. , . – ( ) – . – . , () . , . , . ( – ), . , , , . (” “) , . , . .
, . ( ) , , . , – , – . – , – – , – – (), .
. , . 3 . , .
…
: , . : , .
: 1. , . 2. . 3. . , .. , . , – , . .
, , , . ( – , , , ), (, – , ).
() () – , – ( ). , . – – ( ). , . (), (), – ( – ).
– (), – , – , – .
, . , .
…
. , . , . , , . , . .
(myeloncephalon) . . bulbus cerebri, , I . . (, . ) (, , . ) , .
, , . ( ), , (, , , – ). – , . .
, . IX X , .
(nucleus gracilis et cuneatus) (lemniscus mlis), .
, , ( 100, 102, 105 .). , , . , , (, ).
(n. tractus spinalis nervi trigemini), . .
(metencphalon) : . , – . . , . , . . , . , . , . , – . , , . . : (), . , – , .
( ), ( ) . , – . , , , , . , , . , , , .
(mesencephalon) , . , , . , . : 1) , 2) , 3) . , . , . , , . . , , , , . , , , . , . (), . , , , , , ( ). , , , .
(diencephalon), , , . : thalamencehalon ( ) hypothalamus ( ).
– , , , , , . . 120 . , , . , , , , , , , , . , , , . ( , , , , ) . 3 – 4 . , , . , , . , , . , , , , . , . .
, . , , “” . . , , , – , – . . , . – – (GI4) , , . , . – . . .
() , , , . , . , , , , , , . , , . , . , , , . .
, , . , , , , . – () – (), . , . , , , . , . , , , , .
, , , , . , . .
. () , – , . () , , , , .
. , . (. . , 1983). , , , , . ( , , . ).
( ) : ( ), ( ), . ( , ), , . . .
, , .
, – (, , , ). , , , . , , , , , . , , . , , , . Papez, , , , , , . , , . , , . , . . , , , , . , , , , – , . – . , , .
, , .
. . , , , , , . , . .
. – , , . . . , , . , , , , , , .
( ) . . . , , , . , , . , ( – ). , .
. ; , , ; . , . , 6 – 8 ( 3 – 4 ).
..
( ) (.., 1975). , – – . () .
” ” : . .
, – – . () . . . , . , . , . ( ), , , , , . , . , . – .
…
, , .
. a- 12-22 , 70-120 /, – . , – – ( ). () ( – – ) , . , . ( 4-6 /.). – – , () – . , , – , . . a-. – g-, , , .. .
…
. – – . . , . , . (1) , . , . . – – , . . , .
… – : . , . , , . – .
…
: – . , , , ; . (). – – .. . , , , . , (- ).
, – . , , .. , . , , . .
. ( ), . – , , , , , , , , . .., . , -, .. .
…
(-, -, ) . . , – , . – .
…
, . , .
, . (, , , ), ( , , ) , , . .. (1948).
, , , D, . , (” ” – ..). , ( , .). , , .. – , , , .
, , .. . , – . , , . , . , . . . – , , .
, .. , , .
:
, , ; . 400-700 , ( 300 ), – . . : ( ) () . , “” . . : , (), , . , , ( ), . – (. 1). – – , . , (. 2) (. ). – . – . . (), () 1,8o105 1 ; , . ( 1 ). , ; . . . , – . , ( 2 ).
, , , . – , . , 3 ; . (. ). . , . . – – , , . . ( – 1%) . : (), ( ), , . ( . .), – . , – , . . , . , 15-20 , – 50-60 ( ), (, ) – 250-300 .
…
: , , .. , ( ). , . (“”), , , , . (. 3) . , , , , , ( , ..). , , . . – , ( , , ) , , , . .
–
. , , . , ( ), , ( ) ( ) . (, , ; . ). , – . , , , . (. ) . , , . . .
…
10-20 ( ) 20 ( ). 1-3 ( 2- 10-5 /2). , 140 2- 10-5 /2. (, 50-100 – 3-5 , 100 – 200 ). . (), – , . ( ) 0,3-0,7 , – 2-8 .
…
, , () ( ” ” ). ( , , ) . – , , , , , .
., , . , , . ( ), . (, , ) , . ( ) (. . 1). . , , , ( , ..; . ). …
. , , 1 3 5o10-15 (10 . ). (10 . 1 3) – (100 1 3). , -, 1 . . . , . . , , , ” “. .
…
. , . . , . . , (. 2). , . , . ; . . . , . : ( ) (. 3). , , , . – ” “, (, ).
:
- ..
.
Источник
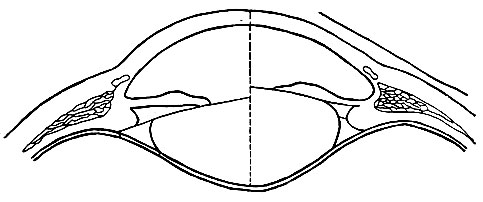
, , . . , . . (), (), , , , , . , . . , , , . . : . , (, , ). , , . . . , , . . . , , . . – – , , . , . . . . . , . – , , . . , . . . . . , , . . – , . , , . , , . , . . , . . () – , 100 . . 59 , – 70,5 . , . , . . , . , . , . , , (. 91). , , . . , , , .
. 91. ( ). – , – . – . , , , (. 92).
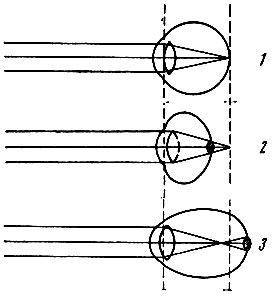
. 92. (1), (2) (3) . , , , . , . . , . , . . , , , . , – . . , , . . . – – . , , , , , . , , – . . , . . . , , , . . . . , 4 . 6-7 . 110-125 . . . ( 140 . 1 2). , . . , , . – () – . . . , , , . 93, ( 25 ) , .
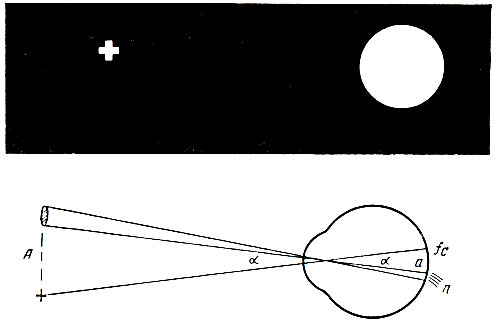
. 93. . . , , (n), – (fc) . , . , , . . , . – . , , , . 17, 18 19- . , – , , . . . . , . . 15 . . , , , . . . . . – , . , , , , . , “” , . , . . . . , , – . . . (1756) , . . . , . , , , – . , , . , , , . . . 620-760 (6200-7600 ) , 510-550 (5100-5500 ) – , 390-450 (3900-4500 ) – . , , , , , . . . , . , . . , , . . . , , , . . . . . , , . . , . . – , , (. 94). – – , . , .
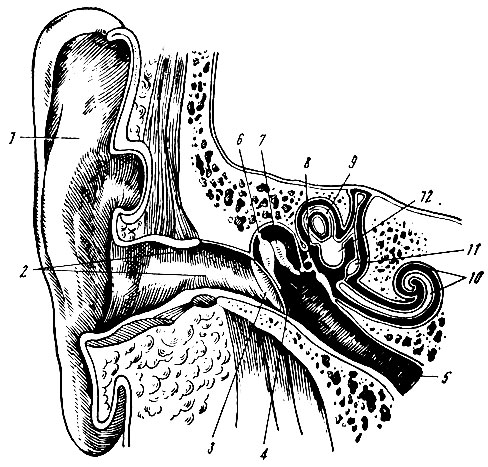
. 94. . 1 – ; 2 – ; 3 – ; 4 – ; 5 – ; 6 – ; 7 – ; 8 – ; 9 – ; 10 – ; 11 – ; 12 – , , , , . , . . , , , , . – () , . , . , , . . () , , , . . , . – , . : , , , , (. 95). . . : , (. . 96).
. 95. ( ). 1 – ; 2 – ; 3 – ; 4 – ; 5 – ; 6 – ; 7 – ; 8 – ; 9 – ; 10 – ; 11 – ; 12 – ; 13 – ,
. 96. . 1, 2, 3 – ; 4 – ; 5, 6 – ; 5 – ; 6 – ( ); 7 – , . . . , . , , . . . . . – . , , . , – . , . . . . . – . . , . : , . . . . , , . . , . , . – 16 20000 1 . . , , . , , . , . – . , . , . , , . . . – , . , , , – , – . (. 96). – () (). , – . , . . . . . , , . . . , . . , . . . . . , , , , , . – . (), (), ( ). () : – , , – , . , , , , , . , , . . . , , , – ( ), – . –, , , , . : , ; , ; , , , . , . , ( ) . . , , -. , , , . . -. , , . , – – , – . , . . . , . . , , , . . ( 2 6) . . – . . . , , , , . . , , . , . , , , – . . , . . , , – . , . . ( ), ( ) , , . , , , . “”, “”, “” . . . , . . , . . , , 40 , . . , . , , . . . . . , , . ,. . , , , . , , , , , . , , . . , , , , , , . , . , . , , . : , , , . , , , . , . , . , – . . : , – . , . , , . , . . , . , , , , , . , , . , . , . , , , XII ( ). . , , . . . , (), (), () (). , . . , , . |

Источник
РЕЦЕПТОРЫ (лат. receptor принимающий) – специализированные чувствительные образования, приспособленные для восприятия адекватных для организма стимулов (раздражителей).
В научной литературе используют также понятие «сенсорные рецепторы» для обозначения Р., обеспечивающих чувствительность (см.) организма. Тем самым разграничиваются сферы применения термина «рецепторы» (в физиологии) и термина «биохимические рецепторы» (используемого в фармакологии, биохимии, иммунологии и др. для обозначения надмолекулярных структур клетки, обеспечивающих взаимодействие с хим. веществами – медиаторами, гормонами и др., и соответствующие реакции клетки).
Сенсорные рецепторы
В относительно просто организованных чувствительных структурах (напр., во внутренних органах, в структурах опорно-двигательного аппарата, в коже) понятие «рецепторы» совпадает с понятиями «рецепторные приборы», «сенсорные органы». В более сложных чувствительных образованиях (напр., в органах слуха и зрения, вестибулярном лабиринте и др.) Р. являются лишь частью сенсорного органа (см. Вестибулярный анализатор, Вкус, Зрение, Осязание, Слух). В Р. энергия раздражителя трансформируется в специфическую активность нервной системы, в сигналы, несущие по афферентным проводникам к нервным центрам информацию о характеристиках действующего агента. В ходе эволюции Р. усложнялись и специализировались, т. к. чем быстрее и полнее организм способен получить информацию о состоянии и об изменениях окружающей и внутренней среды, тем выше шансы организма выжить в непрерывной борьбе за существование.
У высокоорганизованных животных существует большое разнообразие Р., позволяющих им очень точно воспринимать раздражители самых разных видов (модальностей): механические, химические, температурные, световые, электрические. В зависимости от этого различают механорецепторы (см.), хеморецепторы (см.), терморецепторы (см.), фоторецепторы (см.); иногда говорят о наличии ноцицепторов, т. е. рецепторов, воспринимающих болевые раздражители (см. Боль).
Нек-рые рецепторы приспособлены для восприятия одного вида раздражения (мономодальные Р.), другие – для восприятия нескольких видов раздражителей (полимодальные Р.). Исторически сохранилось деление Р. на так наз. дистантные Р., служащие для получения информации на нек-ром расстоянии от источника раздражения, и контактные Р., воспринимающие стимул при непосредственном соприкосновении с ним. Основная масса Р., особенно высокоспециализированных, воспринимает раздражители из окружающих сред. Это так наз. экстероцепторы (см. Экстероцепция). Важную роль играют Р., сигнализирующие о раздражителях внутренней среды, т. е. интероцепторы (см. Интероцепция). Среди них часто выделяют Р. опорно-двигательного аппарата – проприоцепторы (см.).
В зависимости от реакции на длительно действующее стационарное воздействие Р. делят на быстро и медленно адаптирующиеся (фазные и тонические Р.). По структурным и функциональным особенностям Р. подразделяются на первично чувствующие и вторично чувствующие Р. Восприятие стимула в первично чувствующих Р. осуществляется непосредственно (т. е. первично) окончаниями сенсорного нейрона (см. Нервные окончания). У вторично чувствующих Р. между действующим стимулом и сенсорным нейроном располагается специализированная клетка, из к-рой при раздражении выделяется медиатор (см.), действующий уже непосредственно на окончания сенсорного нейрона. Таким образом, у Р. этого типа внешнее раздражение на сенсорный нейрон опосредованно, вторично. К первично чувствующим Р. у позвоночных животных относятся, напр., нервно-мышечные веретена, нервно-сухожильные веретена, а к вторично чувствующим – рецепторы органов слуха, зрения, вкуса, вестибулярного лабиринта и др.
В Р. выделяют три основные части: вспомогательные структуры, напр, капсула у инкапсулированных тканевых Р. (типа телец Пачини, Мейсснера и др.), звукопроводящие структуры органа слуха и др., собственно рецептирующие элементы, содержащие воспринимающий субстрат, и систему генерации локальных электрических потенциалов (так наз. рецепторные, или генераторные, потенциалы). У первично чувствующих Р. ответы возникают в окончании сенсорного нейрона, а у вторично чувствующих Р.- в рецептирующей клетке. Если локальный электрический потенциал оказывает деполяризующее действие на электровозбудимые структуры окончаний сенсорного нейрона, то происходит генерация импульсов (см. Нервный импульс), несущих в ц. н. с. информацию о событиях, протекающих в Р.; отсюда и термин «генераторный потенциал».
Между силой адекватного для Р. раздражения и частотой импульсации (в среднем диапазоне нагрузок) существует логарифмическая зависимость, что соответствует закону Вебера – Фехнера (см. Ощущение), Амплитуда рецепторного потенциала, несмотря на постоянство раздражителя, может флюктуировать. Этот факт, а также флюктуация возбудимости структур, генерирующих импульсы, определяют общее колебание возбудимости Р. во времени, что лежит в основе так наз. функциональной мобильности Р.
Одной из важнейших характеристик Р. является их высокая чувствительность к действию адекватного раздражителя. Чувствительность Р. оценивается величиной абсолютного порога, т. е. минимальной силой раздражения, способной вызвать возбуждение Р. (см. Возбуждение). Абсолютные пороги высокодифференцированных Р. (в органах зрения, слуха, обоняния) могут быть крайне низкими, приближаясь к теоретически предельным значениям.
Деятельность Р. находится под нейрогуморальным контролем. Гуморальные факторы способны в известной мере менять возбудимость Р. Эфферентные нервные влияния могут изменять пороги реакции, вызывать как возбуждение, так и торможение Р. У позвоночных животных эфферентные влияния на высокочувствительные вторично чувствующие Р. носят преимущественно угнетающий характер, а на менее чувствительные первично чувствующие – в основном облегчающий (или возбуждающий) характер.
Патология Р. разнообразна. Причиной могут служить какие-либо нарушения во вспомогательных структурах Р. (напр., в светопроводящих структурах органов зрения, звукопроводящих структурах слуха), в собственно сенсорных элементах (напр., при атрофии обонятельного эпителия, при нарушениях биохим. превращений зрительных пигментов и др.) и, наконец, в афферентных нервных проводниках (напр., при травмах и заболеваниях нервов). Если патол. изменения вспомогательных структур Р. могут подвергаться лечению (напр., при катарактах, отосклерозах), то повреждение собственно рецептирующих элементов (фоторецепторов, волосковых рецепторов и т. д.) обычно ведет к необратимым изменениям в деятельности сенсорных органов.
Методы исследования Р. различны; применяют многие морфологические (свето- и электронно-микроскопические), физиологические (различные микроэлектрофизиологические, психофизиологические и др.), фармакологические, биохимические, биофизические, математические и другие методы.
Клеточные рецепторы
В многоклеточном организме передача информации между клетками, происходящая с участием гормонов, нейротрансмиттеров (медиаторов), нейропептидов и других биологически активных веществ, включает этап взаимодействия молекул этих веществ (их называют также лигандами) с соответствующими надмолекулярными структурами, или клеточными рецепторами. Они могут располагаться как внутри клетки (напр., клеточные Р. к стероидным гормонам, легко проникающим внутрь клетки благодаря их растворимости в липидах клеточной мембраны), так и на поверхности клеточной мембраны (клеточные Р. к белкам, пептидам, нейротрансмиттерам). Как внутриклеточные, так и мембранные клеточные Р. содержат центр связывания, обеспечивающий специфическое связывание лиганда с клеточными Р. После связывания, напр., молекулы стероидного гормона с цитоплазматическим Р. и образования комплекса гормон – клеточный Р. этот комплекс проникает внутрь клеточного ядра, где связывается с соответствующим акцептором, вслед за чем молекула гормона отделяется от комплекса и выходит в цитоплазму, при этом одновременно активируется генетический аппарат клетки (см.). Конечным итогом этой активации является резкое усиление синтеза ряда специфических и неспецифических белков клетки, что представляет собой ответ клетки-мишени на действие гормона.
Процессы, происходящие при связывании молекулы лиганда с клеточным Р., локализованным на клеточной мембране (см. Мембраны биологические), состоят из ряда этапов, протекающих с большой скоростью. Происходящее при этом изменение свойств фосфолипидного матрикса, окружающего клеточный Р., обеспечивает передачу сигнала от центра связывания лиганда (через ряд промежуточных звеньев) на аденилат-циклазный центр и его активацию. Циклическая АМФ (см. Аденозинфосфорные кислоты) является своеобразным вторым внутриклеточным переносчиком информации, определяющим ответ клетки на действие лиганда. Т. о., происходит активация соответствующих протеинкиназ, изменение проницаемости клеточной мембраны для ряда ионов, усиление экспрессии генетической информации. Важным открытием явилось обнаружение в ц. н. с. клеточных Р. к ряду нейрональных пептидов, напр, к группам пептидов, названных эндорфинами и энкефалинами, а также клеточных Р. ко многим психотропным лекарственным препаратам (имипрамину, галоперидолу, диазепаму и др.). Взаимодействие указанных выше лигандов с клеточными Р. изменяет способность нервных клеток отвечать на действие нейро-трансмиттеров, т. е. оказывает на их активность модулирующее действие. Напр., связывание препаратов бензодиазепинового ряда с бензодиазе-пиновыми клеточными Р. усиливает ответ ГАМК-ергических нейронов на действие гамма-аминомасляной к-ты (ГАМК), влияя в то же время на связывание ГАМК соответствующими клеточными Р. В связи с обнаружением в ц. н. с. эндогенных лигандов, конкурирующих с морфином за места связывания и обладающих морфиноподобным действием, проводится поиск эндогенных соединений типа эндогенного диазепама, эндогенного галоперидола и др., что может иметь в случае их обнаружения большое значение для клин, практики.
Нарушения механизмов рецепции играют важную роль в развитии ряда заболеваний человека, напр, нек-рых видов сахарного диабета, гиперхоли-стеринемии и др. Наряду с рассмотренными выше видами Р. на поверхности В- и Т-лимфоцитов выявлены мембранные клеточные Р., играющие важную роль в работе иммунной системы, а также клеточные Р. к ряду вирусов.
Библиография: Глебов Р. И. и Крыжановский Г. Н. Функциональная биохимия синапсов, М., 1978; Гранит Р. Электрофизиологическое исследование рецепции, пер. с англ., М., 1957; Розен В. Б. и Смирнов А. Н. Рецепторы и стероидные гормоны, М., 1981, библиогр.; Тамар Г. Основы сенсорной физиологии, пер. с англ., М., 1976; Физиология сенсорных систем, под ред. А. С. Батуева, с. 34, Л., 1976; Cell membrane receptors for drugs and hormones, a multidisciplinary approach, ed. by R. W. Straub a. L. Bolis, N. Y., 1978; Cell! membrane receptors for viruses, antigens, and antibodies, polypeptide hormones, and small molecules, ed. by R. F. Beers a. E. G. Bassett, N. Y., 1976; The receptors, a comprehensive treatise, ed. by R. D. O’Brien, v. 1, N. Y.- L., 1979.
О. Б. Ильинский; P. P. Лидеман (клеточные рецепторы).
Источник