
Сердце с сосудами ящерица

Сердце ящерицы окружено околосердечной оболочкой и состоит из крайне редуцированной, наполовину слившейся с правым предсердием венозной пазухи, правого и левого предсердий и одного, не вполне разделенного перегородкой желудочка. Тонкостенная венозная пазуха, в которую впадают крупные вены, сообщается с правым предсердием, отверстие в которое защищено двугубым клапаном. Предсердия открываются в желудочек вытянутым отверстием, поделенным надвое перегородкой предсердий и снабженным клапаном из двух полулунных складок. Таким образом, для каждого предсердия диференцируется свое отверстие в желудочек, снабженное двумя клапанами. Это представляет уже дальнейшую стадию развития и диференциации сравнительно с амфибиями.
Относительно небольшая полость желудка, несущая внутримышечные складки, причем мышцы подтягиваются сухожилиями (chordae tendineae), разделена надвое неполной перегородкой. Эта перегородка начинается на нижнебрюшной части желудочка, наклонена слева направоы делит желудок таким образом, что получаются левое спинно-боковое отделение общего желудочка и правое вентрально-боковое отделение. Когда происходит систола (сокращение) желудочка, то дорзальные стенки его соприкасаются с соответствующим краем перегородки и на момент осуществляется, правда, неполное разделение двух половин желудочка.
Рис. 1. Схема кровообращения ящерицы
1—общий ствол сонных артерий (a. carotis communis); 2—внутренняя сонная артерия; 3—наружная сонная артерия; 4—легочная артерия; 5—левая дуга аорты; 6—правая дуга аорты; 7—подключичная артерия; 8—легочная вена; 9—полая вена (нижняя) и две яремные вены (верхние); 10— спинная аорта; 11—arteria coeliaca (к внутренностям); 12 — ductus caroticus; 13 — ductus botalli (не развитый).
Прогрессивным признаком строения сердца ящерицы и прочих рептилий надо считатъ полное исчезновение артериального конуса. Ток крови от сердца направляется по трем главным путям—по двум дугам аорты—правой и левой (рис. 2 , 5—6) и по легочной артерии (рис. 2 , 4), которая самостоятельным стволом начинается от правой части желудочка. Что касается дуг аорты, то дуга, поворачивающая направо, у всех рептилий начинается особым стволом, берущим начало в левой части желудочка (рис. 2 , 6). От правого ствола берут начало сонные артерии, имеющие общий ствол (arteria carotis communis; рис. 2 , 1); на дальнейшем протяжении этот общий ствол разделяется на внутреннюю и внешнюю сонные артерии (a. carotis interna и а. с. externa; рис. 2 , 2—3). У ящериц каждая внешняя сонная артерия иногда сообщается тонким протоком (ductus caroticus) с соответствующей дугой аорты (рис. 2 , 12).

Рис. 2 — 3. Сердце ящерицы. Сердце черепахи.
Обозначения 1—9, как на предыдущем рисунке. 8 — яремные вены; 10—11, как на предыдущем рисунке; 12—боталлов проток.
От правой дуги аорты берут начало подключичные артерии. У черепахи они начинаются непосредственно от соответствующих сонных артерий (рис. 3 , 7).
Левая дуга аорты у черепахи начинается от правого отдела желудка (рис. 3 , 5), у ящериц—на границе между правым и левым отделами, примерно против перегородки между частями желудочков (рис. 2 ). Она делает изгиб и дорзально соединяется с правой дугой, образуя д о р з а л ь н у ю, или спинную, аорту или артерию (рис. 3 , 10). От левой дуги у всех рептилий берет началоартерия, отходящая к внутренним органам (arteria coeliaca; рис. 2 , 3 , 11).
Венозная кровь возвращается из хвостового отдела по хвостовой вене (рис. 5 , 19), которая близ основания хвоста распадается на две тазовые вены; соединившись, тазовые вены образуют брюшную вену (рис. 5 , 21), идущую в левую лопасть печени. В тазовые вены изливают кровь вены, идущие от задних конечностей; от тазовых же вен берут начало две воротные вены почек, дающие разветвленную воротную систему в вещество почек. Выносящие почечные вены (venae renales revehentes) вскоре сливаются в заднюю полую вену (рис. 5 , 24); на своем пути к сердцу этот крупный кровеносный сосуд принимает большую печеночную вену и впадает в едва заметную (почти редуцированную) венозную пазуху, куда приносят кровь от головы и передних конечностей две передние полые вены (рис. 5 , 28).

Рис. 4 . Схема строения сердца рептилий
I—продольный разрез сердца; II—поперечный разрез сердца черепахи, III — сердца ящерицы. 1 — левое предсердие; 2 — правое предсердие; 3 — желудочек; 4 — перегородка в желудочке; 5—легочная артерия; 6—левая дуга аорты; 7—правая дуга аорты.
Печень снабжена воротной системой сосудов, причем ток венозной крови приносится в нее воротной веной, образующейся от соединения брюшной вены, вены поджелудочной железы и брыжейки (рис. 5 , 25).
В левое предсердие поступает кровь через легочные вены, которые перед тем, как войти в предсердие, сливаются в общий ствол.
Ознакомившись со строением сердца и отходящих от него сосудов, а также сосудов, приносящих кровь к сердцу, легко представить себе последовательность процесса кровообращения. Венозная кровь поступает в желудочек из правого предсердия, артериальная (окисленная)—из левого. Несомненно, что эти два потока крови в желудочке смешиваются. Однако это частично устраняется и затрудняется наличием, хотя и неполной, перегородки, а также рядом мелких подразделений, имеющихся в нижней части желудка и разгороженных мышцами.
Рис. 5. Кровеносная система ящерицы
Без сомнения, ствол аорты, поворачивающийся налево, берущий начало в сердце черепахи в правой части желудочка, захватывает венозную кровь (с очень малой примесью артериальной). У ящерицы, как мы видели, этот ствол отнесен дорзально и расположен ближе к средней части желудочка, вследствие этого здесь примешивается больше артериальной крови, чем у черепах. Легочная артерия получает венозную кровь и несет ее для окисления в легкие (по малому кругу кровообращения). Дуга аорты, поворачивающаяся направо, у всех рептилий берет начало в левой части желудочка. В ее кровяном токе, естественно, преобладает артериальная кровь, идущая к голове (по сонным артериям) и к конечностям (по подключичным артериям). В спинной аорте (артерии) смешивается кровь правого и левого стволов аорты, т. е. циркулирует кровь смешанного типа. Это, без сомнения, указывает на менее совершенное строение системы кровообращения у рептилий, чем у птиц и млекопитающих.
Это несовершенство не устраняется и в кровеносной системе крокодила, несмотря на полное разделение желудочка перегородкой. В общее кровяное русло левый ствол аорты и у крокодила доставляет примесь венозной крови.
У черепах и у гаттерии каждая легочная артерия соединяется узким протоком (ductus Botalli) с соответствующей дугой аорты (рис. 2 ,13).
Статья на тему Органы кровообращения ящерицы
Источник
Васильев Дмитрий Борисович
врач-герпетолог
Особенности строения и работы сердечно-сосудистой системы ящериц
Остановимся только на нескольких «узловых» моментах, отличающих ящериц от высших позвоночных.
- Сердце у ящериц относительно небольших размеров.
У рептилий масса сердца составляет около 0,2–0,3% от массы тела, что больше, чем у рыб, но меньше, чем у амфибий. - Положение сердца варьирует у многих рептилий.
У ящериц с вытянутым телом, особенно, у варанов, сердце располагается за рукояткой грудины, то есть почти на середине тела. - Частота сердечных сокращений (ЧСС) у рептилий обратно пропорциональна массе тела (как у всех позвоночных) и прямо пропорциональна температуре тела и окружающей среды (как у всех холоднокровных).
У ящерицы массой 1 кг ЧСС должна составлять около 33 ударов/мин. На самом деле у игуан даже при температуре 24°С ЧСС обычно составляет не менее 40–70 ударов/мин (Bennett, Schumacher, et al, 1998) и только у очень крупных ящериц она может быть меньше 30 ударов/мин. - Все рептилии, за исключением крокодилов, имеют трехкамерное сердце с неполной межжелудочковой перегородкой, и поэтому в большей части кровеносной системы кровь остается смешенной.
- Трехкамерное сердце рептилий «технически» позволяет регулировать распределение сердечного выброса между большим и малым кругом кровообращения с помощью интервентрикулярного шунта (преимущественной подачи потока крови в правый или левый желудочек сердца).
Многие рептилии способны к периодической задержке дыхания. При этом сосудистое сопротивление в легких возрастает, давление в малом круге повышается, и кровь шунтируется (сбрасывается) в левое сердце и в большой круг — наблюдается левый шунт, который позволяет выключать циркуляцию в малом круге в состоянии апноэ (отсутствия дыхания). И, наоборот, в процессе самостоятельного дыхания преобладает правый шунт. - Рептилии способны к анаэробному гликолизу (процесс распада глюкозы без участия кислорода с образованием молочной кислоты), хотя это видоспецифично.
Некоторые ящерицы могут выживать без кислорода не более 25 минут, а пресноводные черепахи — более 33 часов. Игуаны выживают в бескислородной среде около 4,5 часов (Moberly, 1968). Это зависит в первую очередь от устойчивости миокарда к гипоксии (пониженному содержанию кислорода в его клетках). При этом сердечный выброс уменьшается до 5% от нормы, и кровь направляется в основном к голове и главным внутренним органам. Это позволяет рептилиям поддерживать только жизненно важные функции, в отличие от ныряющих птиц и млекопитающих.
При нормальном воздухообмене уровень сердечной деятельности мгновенно восстанавливается. - Рептилии, подобно птицам, рыбам и амфибиям имеют воротную систему почек.
Кровь туда собирается из капиллярного ложа задних конечностей и хвоста. Конечно, под воздействием многих факторов, кровоток может изменять своё направление и обходить воротную систему почек по анастомозам с брюшной веной. Это происходит, прежде всего, при изменении температуры тела и статуса гидратации. Но, при возможности выбора лучше инъецировать препараты в переднюю половину тела, чтобы избежать их преждевременного выделения почками и нефротоксического действия некоторых препаратов при увеличении их концентрации в воротной системе почек. - У ящериц по белой линии живота примерно в середине тела непосредственно под брюшной мускулатурой расположена крупная вентральная брюшная вена.
Поэтому, уже давно при полостных операциях у ящериц выбирают парамедианный доступ (т. е. продольный разрез делают не по центральной линии живота, а в стороне от неё), чтобы избежать травмирования вены и кровотечения. Но всё-таки хирургический доступ по белой линии имеет свои преимущества: доступ равнозначен для парных органов, кроме того, он уменьшает постоперационную боль и дискомфорт, а также при этом не приходится пересекать мышцы брюшной стенки. У крупных видов ящериц, брюшную вену можно сдвинуть вбок от белой линии в любом направлении. В случае пересечения и/или лигирования (перевязки) брюшной вены кровь оттекает обратно через тазовые и почечные вены в заднюю полую вену. Поэтому выключение данного сосуда из кровотока не вызывает серьезных нарушений циркуляции крови. Мы редко используем доступ по белой линии, так как в этом случае швы находятся в постоянном контакте с грунтом. Однако, это удобный и быстрый доступ к брюшной вене для её катетеризации и, как менее кровавый, оптимален для пациентов с возможной коагулопатией.
Источник: ветклиника «Белый Клык»
Источник

Еще фото
Автор (ы):
Л.А. Стоянов, врач ветеринарной медицины, начальник ветеринарно-медицинского отдела экзотических животных Международной ассоциации океанариумов и дельфинариев
Организация(и):
Сеть океанариумов «Немо», г. Одесса
Журнал:
№2 – 2013
Окончание. Начало в номере №1 за 2013 год
Благодарим редакцию журнала «Мир ветеринарии», Украина, за любезно предоставленную статью Л.А. Стоянова
Анатомия и физиология дыхательной системы
Знание нормальной анатомии и физиологии дыхательных систем рептилий крайне важно для врачебной практики. Хотя общее строение и функции легких у разных отрядов рептилий в целом одинаковы, у отдельных групп были найдены некоторые клинически значимые отличия. Недостаточное понимание анатомии и физиологии дыхательной системы рептилий не позволит эффективно лечить такие заболевания, как пневмония, и отрицательно скажется на умении обращаться с рептилией под действием анестезии.
Анатомия дыхательных путей
Дыхательные системы рептилий и млекопитающих сильно отличаются анатомически и физиологически. Существенные отличия наблюдаются также у разных отрядов рептилий и между видами внутри одного отряда, особенно при сравнении водных и наземных видов. Ниже приведены основные клинически значимые особенности анатомии дыхательных систем черепах, ящериц, крокодилов и змей.
Черепахи
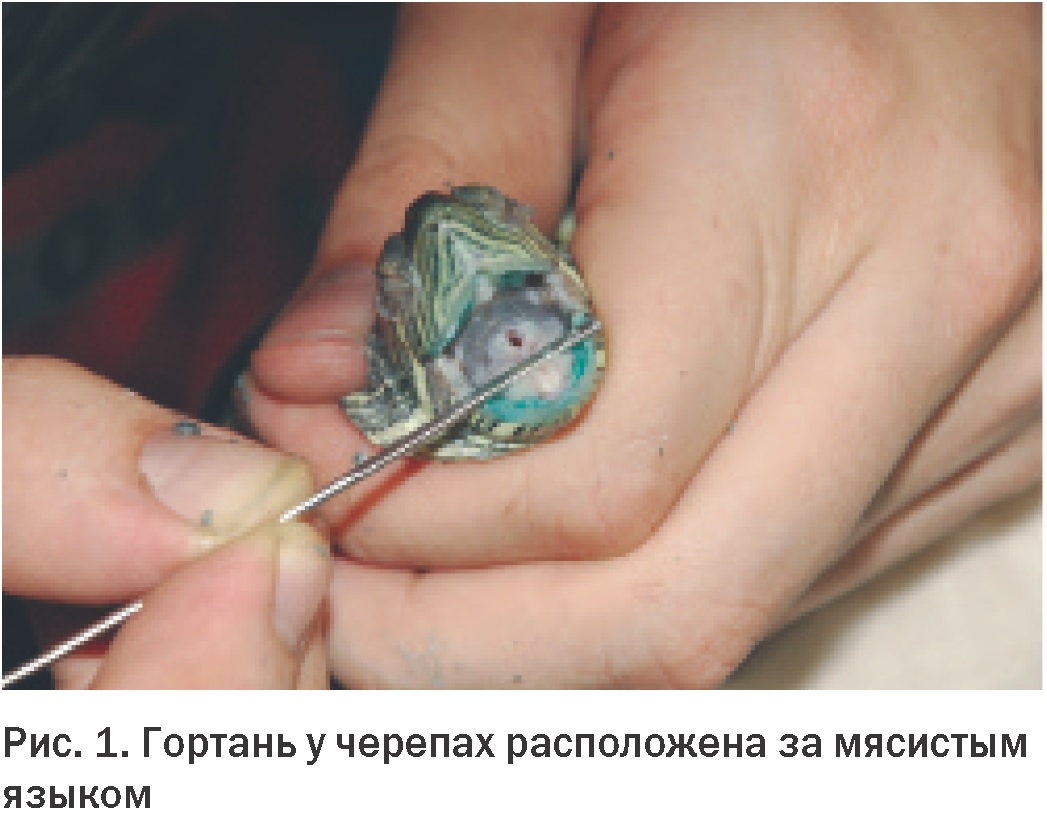
Гортань у черепах открывается у основания толстого мышечного языка в относительно каудальной части ротоглотки (Рис. 1). Такое расположение затрудняет доступ к нижним дыхательным путям через ротовую полость без применения наркоза. Трахея с полными хрящевыми кольцами раздваивается чуть ниже шеи. Далее парные бронхи в жесткой оболочке входят в легкое с дорсальной стороны.
Легкие черепах парные и мешковидные. Структуры, в которых происходит газообмен, названные фальвеолами, в противоположность альвеолам млекопитающих, открываются на окончаниях бронхов в воздушную полость легкого. При ближайшем рассмотрении структуры легкого обнаруживаются внутренние выступы и перегородки, напоминающие губку в поперечном разрезе. Общая анатомия нижних дыхательных путей черепах имеет большое значение. Воспалительный экссудат, в частности, образующийся при инфекционных заболеваниях, имеет свойство скапливаться в нижней части легкого, что препятствует его высвобождению через бронхи и трахею, как это происходит у млекопитающих.
У черепах прохождение газов через дыхательную поверхность для газообмена достигается различными путями. Эти различия зависят от вида, а также от того, водным или наземным он является. Как правило, и вдох, и выдох являются активными процессами. Следует, однако, отметить, что у водных видов выдыхание является более пассивным, тогда как у некоторых наземных видов более пассивным является вдох. Несмотря на то, что у черепах нет настоящей диафрагмы, многие из них обладают подобной ей мембранной структурой, которая частично разделяет грудную и брюшную полости и называется septum horizontale. У морских черепах это разделение отсутствует.
Как правило, у водных черепах в респираторном цикле работают четыре группы мышц. Сокращение диафрагмальных и поперечных мышц живота сжимает целомическую полость, что приводит к выдоху. Коракоидные и косые мышцы живота расширяют полость и обеспечивают вдох.
Интересно, что повторяющиеся горловые движения, которые владельцы животных часто принимают за дыхание, в действительности с ним не связаны и имеют обонятельную функцию. У сухопутных средиземноморских черепах (Testudo graeca) были найдены существенные отличия. У этого вида дыхательные движения обеспечивают мышцы плечевого пояса. Вдох происходит с сокращением зубчатых и косых мышц живота, а выдох – с сокращением поперечных мышц живота и грудных мышц.
Ящерицы
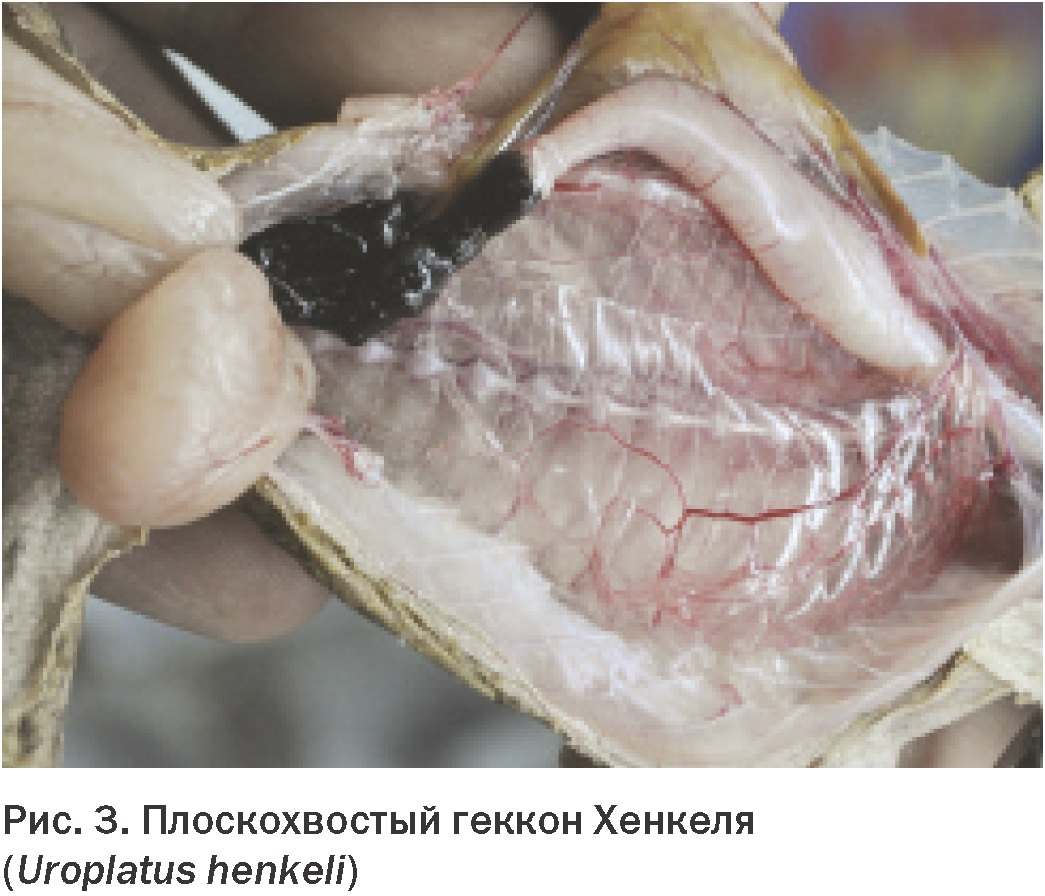
У ящериц расположение отверстия гортани в ротовой полости неодинаково (Рис. 2). У многих видов, в частности, у хищных, оно находится в ростральной части рта. У других – оно расположено вентральнее, у основания мясистого языка. У ящериц, как и у змей, кольца трахеи неполные. В отличие от черепах, их трахея не раздваивается в шейном отделе, а остается цельной до входа в грудную полость около сердца. Ниже трахея делится на левый и правый бронхи. Легкие ящериц примерно одинаковы по размеру и объему и их строение такое же, как у других рептилий (Рис. 3). У многих ящериц газообмен происходит в краниальной части легкого. Каудальное расширение легкого аналогично воздушному мешку у птиц. Эта особенность важна, так как продукты воспалительных процессов могут накапливаться в этой нереспираторной части легкого. В связи со слабой васкуляризацией этой области, инфекции могут удерживаться здесь и не поддаваться действию парентеральных медикаментов.
Как и у черепах, вдохи и выдохи у ящериц являются активными процессами. У большинства видов они совершаются при помощи межреберных мышц. Некоторые виды используют также гладкую мускулатуру внутри легкого для вдоха. У большинства видов сразу за вдохом и выдохом следует период задержки дыхания, различный по продолжительности. У некоторых ящериц, например, у варанов есть мембрана, отделяющая сердце и легкие от остальных внутренних органов, аналогично диафрагме между грудной и брюшной полостями у млекопитающих. Эта мембрана не имеет дыхательной функции.
Змеи
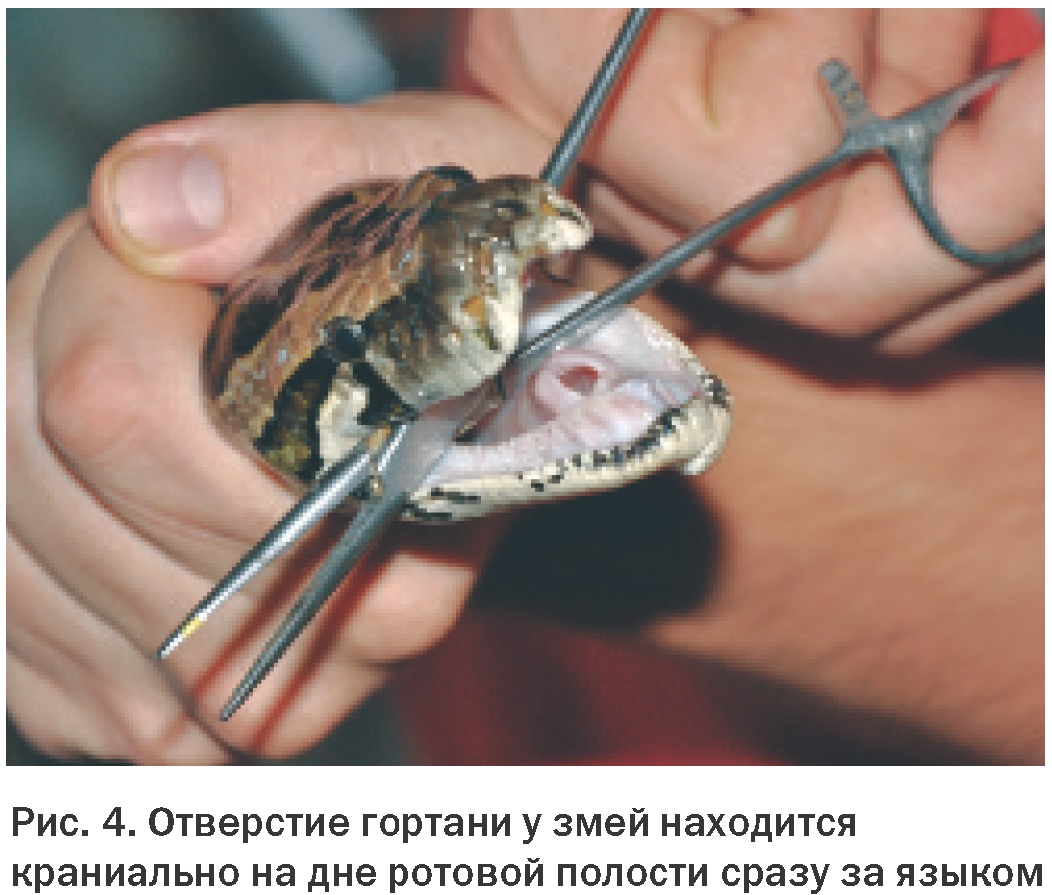
Отверстие гортани у змей расположено ближе к носу в ротовой полости, позволяя таким образом дышать во время заглатывания пищи (Рис. 4). Это также делает возможным прямой осмотр и интубацию трахеи у большинства змей без применения наркоза. Трахея с неполными С-образными кольцами входит в легкое на уровне нижней части сердца. У некоторых видов есть «трахейное легкое», расположенное на дорсальной стороне трахеи. Эта структура служит для продолжения газообмена в случае сдавливания легких проглоченной добычей.
У большинства змей левое легкое рудиментарно или отсутствует. У самых примитивных видов, таких как удавы, есть оба легких, но левое меньше. В легких большинства змей различают два отдела. Первая треть или половина легкого функционирует нормально, а задняя часть относительно мало пронизана кровеносными сосудами и похожа на воздушный мешок у птиц. Краниальная или дыхательная часть легкого обычно начинается на уровне 20% и заканчивается на уровне 40% от общей длины от носа до клоаки. Каудальная или мембранная часть легкого располагается далее и имеет разную длину у разных видов.
Дыхательный цикл змей включает активную и пассивную стадии. Выдох контролируется дорсолатеральным (поперечная мышца спины и верхняя внутренняя реберная мышца) и вентролатеральным (поперечная и косая мышцы живота) слоями мышц. Расслабление мышц, обеспечивающих выдох, является первым пассивным этапом вдоха. Подъём ребер в результате сокращения мышц, поднимающих ребро, и мышц, оттягивающих ребро, является активной частью вдоха. Последняя стадия дыхательного цикла, пассивный выдох, происходит в результате расслабления мышц вдоха и спадания легкого.
Крокодилы
Гортань у крокодилов открывается каудально за хорошо выраженным надгортанным клапаном. Вместе с подъязычной костью он может запирать ротовую полость, позволяя животному дышать при частичном погружении даже с открытым ртом. Полость внутренних органов у них разделена на две части мышечной перегородкой, аналогичной диафрагме. К ней примыкает краниальная часть печени. Каудально печень связана с тазовым поясом парой диафрагмальных мышц.
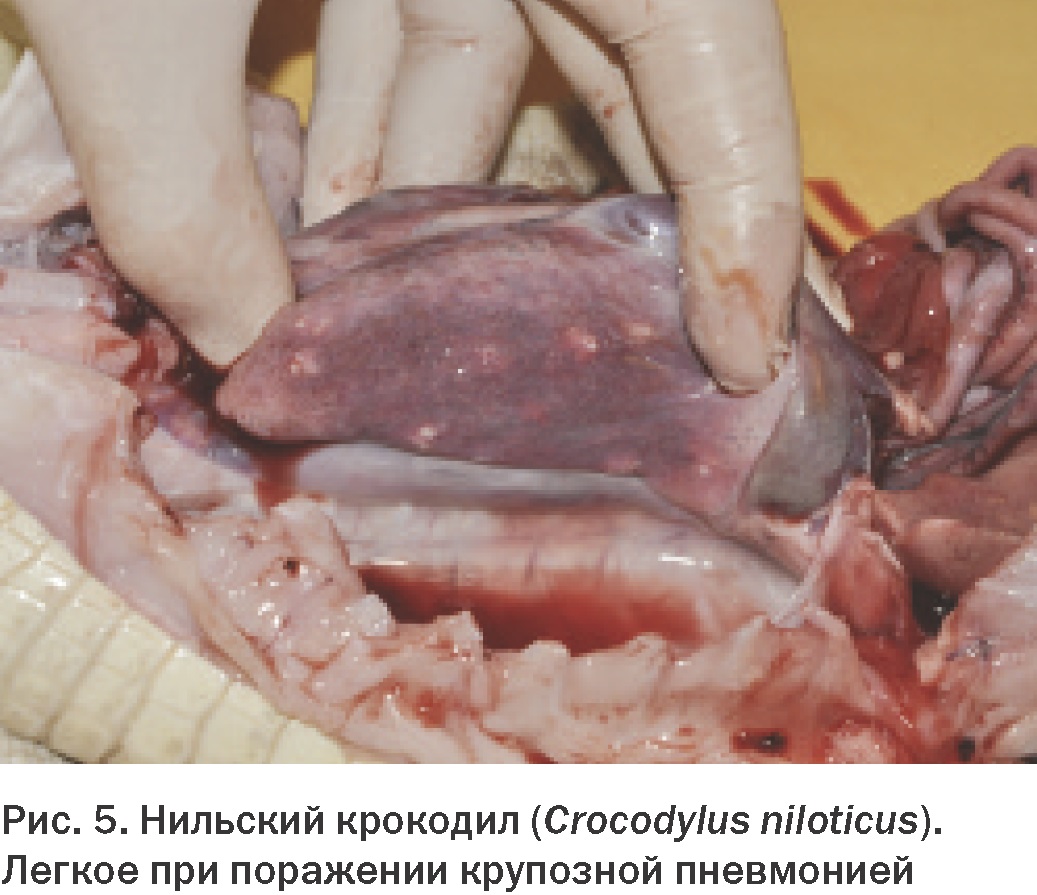
Сокращение этих мышц совместно с межреберными мышцами также направлено на вдох и часто называется «гепатическим поршнем». Активный выдох производится благодаря сжатию полости тела поперечными мышцами. Легкие крокодилов устроены сложнее, чем у других рептилий. Парабронхи делят их на ряд отдельных камер (Рис. 5).
Несмотря на огромные различия в общей анатомии дыхательной системы рептилий, ее микроскопическое строение довольно единообразно. Трахею выстилает многорядный, мерцательный, цилиндрический эпителий с различным количеством бокаловидных клеток. По окончанию крупных дыхательных путей большого диаметра мерцательный эпителий исчезает и заменяется чешуйчатым. Сетчатое альвеолярное легкое состоит из тонкостенных капилляров, проходящих между эпителиальными стенками фальвеол. Каудальное легкое без кровеносных сосудов выстлано простым чешуйчатым эпителием.
Физиология дыхания
Говоря о физиологии дыхания, следует рассмотреть три составляющие респираторного механизма: во-первых, движение респираторных газов из внешней среды к поверхности газообмена, во-вторых, перемещение кислорода и углекислого газа сквозь дыхательный эпителий и, в-третьих, циркуляцию крови, несущей кислород и углекислый газ, от легких к тканям и последующую диффузию газов на клеточном уровне.
Как было показано ранее, рептилии обладают уникальными системами ассимиляции и удаления респираторных газов. Хотя способы работы этих систем весьма вариабельны у разных видов рептилий, все остальные дыхательные процессы в целом одинаковы.
Важное клиническое значение имеет способность рептилий к анаэробному метаболизму. Особые буферные системы, присутствующие в кровеносной системе, позволяют управлять накоплением молочной кислоты и ионов водорода в периоды анаэробиоза. Из-за этой способности даже тяжелые заболевания дыхательной системы могут оставаться скрытыми, пока их прогрессия не превзойдет компенсаторные свойства организма.
Газообмен у рептилий не ограничивается легочными фальвеолами. Многие водные виды способны к газообмену через покровы. Такие черепахи, как мягкотелая черепаха (Apalone spp.), могут получать до 70% кислорода во время погружения через кожистый панцирь.
Щёчно-глоточные перепонки также являются поверхностями для газообмена у множества рептилий, а густо пронизанные кровеносными сосудами глоточные папиллы способны извлекать кислород, растворенный в воде. Мягкотелые черепахи могут получать достаточное количество кислорода через кожу и ротоглотку во время длительных погружений, например, во время спячки.
Многие виды ящериц также способны в некоторой степени к глоточному и кожному дыханию.
Как было упомянуто ранее, некоторые виды змей отличаются особым мешковидным выпячиванием открытых колец трахеи, которое служит для газообмена в том случае, если содержимое желудка мешает нормальной работе легкого. Дыхание пойкилотермных животных контролируется парциальным давлением кислорода, углекислого газа и температурой. При увеличении температуры потребность в кислороде также возрастает. Она обычно компенсируется не учащением дыхания, а увеличением дыхательного объема. Более того, гиперкапния (увеличение содержания CO2) также вызывает увеличение дыхательного объема, тогда как гипоксия приводит к учащению дыхания. Стимулом к учащенному дыханию у рептилий является низкая концентрация кислорода в крови.
При значительной легочной патологии способность пациента к увеличению дыхательного объема ухудшается из-за воспалительных процессов, клеточной инфильтрации и потери эластичности тканей.
Клинически важно, что условия с большим содержанием кислорода в воздухе ведут к подавлению собственных дыхательных движений у рептилий. Хотя отсутствие диафрагмы и слабый бронхоцилиарный транспорт ухудшает отход воспалительного экссудата из легких, длительная экспозиция в насыщенной кислородом среде еще более затруднит его в связи с подавлением вентиляции. Поэтому использование кислородных камер следует применять только в случаях подозрения на гипоксемию. Относительная неэффективность легких в сочетании со способностью рептилий переключаться на анаэробный тип обмена веществ играют важную роль при патологиях дыхательной системы.
Серьезные респираторные заболевания могут вызывать увеличение сопротивления легочных кровеносных сосудов. Так как эти изменения сходны с таковыми при задержке дыхания или погружении, они ведут к изменениям в работе сердца. При нормальном дыхании сердце рептилий, кроме крокодилов, направляет кровь, наиболее бедную кислородом, в легочную артерию. Увеличение сопротивления легочных сосудов приводит к отводу крови от легких в системный круг кровообращения. Если отвод возникает в результате патологии, перенос кислорода к периферическим тканям может быть затруднен. При продолжительном анаэробиозе в результате перегрузки буферной системы крови развивается ацидоз, представляющий угрозу для жизни. ■
Литература
1. Jackson CG Jr: Cardiovascular system. In Harless M, Morlock H, editors: Turtles: perspectives and research, Melbourne, Fla, 1979, Krieger Publishing.
2. White FN: Circulation. In Gans C, editor: Biology of the reptilia, vol 5, New York, 1976, Academic Press.
3. Williams DL: Cardiovascular system. In Beynon PH, Law-ton MPC, Cooper JE, editors: Manual of reptiles, Cheltenham, United Kingdom, 1992, British Small Animal Veterinary Association.
4. Smith JA, McGuire NC, Mitchell MA: Cardiopulmonary physiology and anesthesia in crocodilians, Proc Assoc Reptil Amphib Vet 17-21, 1998.
5. Holz PH: The reptilian renal portal system — a review, Bull Assn Reptil Amphib Vet 9(1):4-9, 1999.
6. Davies PMC: Anatomy and physiology. In Cooper JE, Jackson OF, editors: Diseases of the Reptilia, vol 1, New York, 1981, Academic Press.
7. Anderson JB, Rourke BC, Caiozzo VJ, Bennett AF, Hicks JW: Postprandial cardiac hypertrophy in pythons, Nature (434): 37-38, 2005.
Назад в раздел
Источник