
Слой яичника в котором проходят сосуды и нервы

Сосуды и нервы яичника
Кровоснабжение яичников осуществляется за счет аа. et vv. ovaricae et uterinae. Обе яичниковые артерии (аа. ovaricae dextra et sinistra) отходят от передней поверхности аорты чуть ниже почечных артерий, правая чаще берет начало от аорты, а левая – от почечной артерии. Направляясь вниз и латерально по передней поверхности большой поясничной мышцы, каждая яичниковая артерия пересекает спереди мочеточник (отдавая ему веточки), наружные подвздошные сосуды, пограничную линию и вступает в полость таза, располагаясь здесь в подвешивающей связке яичника. Следуя в медиальном направлении, яичниковая артерия проходит между листками широкой связки матки под маточной трубой, отдавая ей ветви, и далее – в брыжейку яичника; вступает в ворота яичника.
Ветви яичниковой артерии широко анастомозируют с яичниковыми ветвями маточной артерии. Венозный отток из яичников осуществляется прежде всего в яичниковое венозное сплетение, располагающееся в области ворот яичника. Отсюда отток крови проходит по двум направлениям: через маточные и яичниковые вены. Правая яичниковая вена имеет клапаны и впадает в нижнюю полую вену. Левая яичниковая вена впадает в левую почечную вену, причем клапаны в ней отсутствуют.
Лимфоотток от яичников происходит через лимфатические сосуды, особенно обильно в области ворот органа, где выделяют подъяичниковое лимфатическое сплетение. Затем лимфа отводится по ходу яичниковых лимфатических сосудов к парааортальным лимфатическим узлам.
Иннервация яичников
Симпатическая – обеспечивается постганглионарными волокнами из чревного (солнечного), верхнебрыжеечного и подчревного сплетений; парасимпатическая – за счет внутренностных крестцовых нервов.
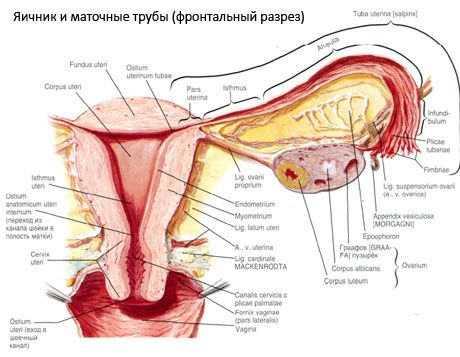
Строение яичника
Поверхность яичника покрыта однослойным зародышевым эпителием. Под ним залегает плотная соединительнотканная белочная оболочка (tunica albuginea). Соединительная ткань яичника образует его строму (stroma ovarii), богатую эластическими волокнами. Вещество яичника, его паренхиму, делят на наружный и внутренний слои. Внутренний слой, лежащий в центре яичника, ближе к его воротам, называют мозговым веществом (medulla ovarii). В этом слое в рыхлой соединительной ткани располагаются многочисленные кровеносные и лимфатические сосуды и нервы. Наружный слой яичника – корковое вещество (cortex ovarii) более плотный. В нем много соединительной ткани, в которой располагаются созревающие первичные фолликулы яичника (folliculi ovarici primarii), вторичные (пузырчатые) фолликулы (folliculi ovarici secundarii, s.vesiculosi), a также зрелые фолликулы, граафовы пузырьки (folliculi ovarici maturis), а также желтые и атретические тела.
В каждом фолликуле находится женская половая яйцеклетка, или овоцит (ovocytus). Яйцеклетка диаметром до 150 мкм, округлая, содержит ядро, большое количество цитоплазмы, в которой, помимо клеточных органелл, имеются белково-липидные включения (желток), гликоген, необходимые для питания яйцеклетки. Свой запас питательных веществ яйцеклетка обычно расходует в течение 12-24 ч после овуляции. Если оплодотворение не наступает, яйцеклетка погибает.
Яйцеклетка человека имеет две покрывающие ее оболочки. Кнутри находится цитолемма, которая является цитоплазматической мембраной яйцеклетки. Снаружи от цитолеммы располагается слой так называемых фолликулярных клеток, защищающих яйцеклетку и выполняющих гормонообразующую функцию – выделяют эстрогены.
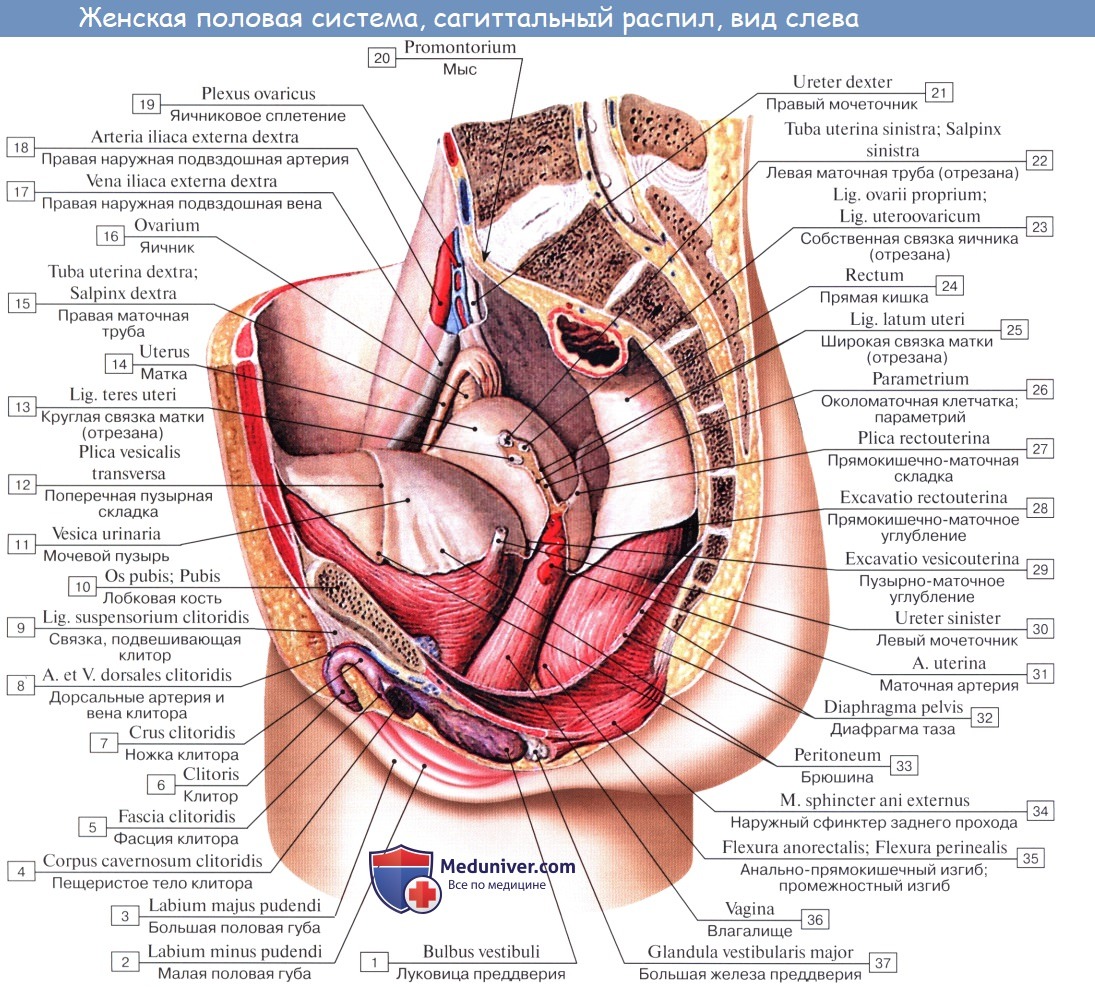
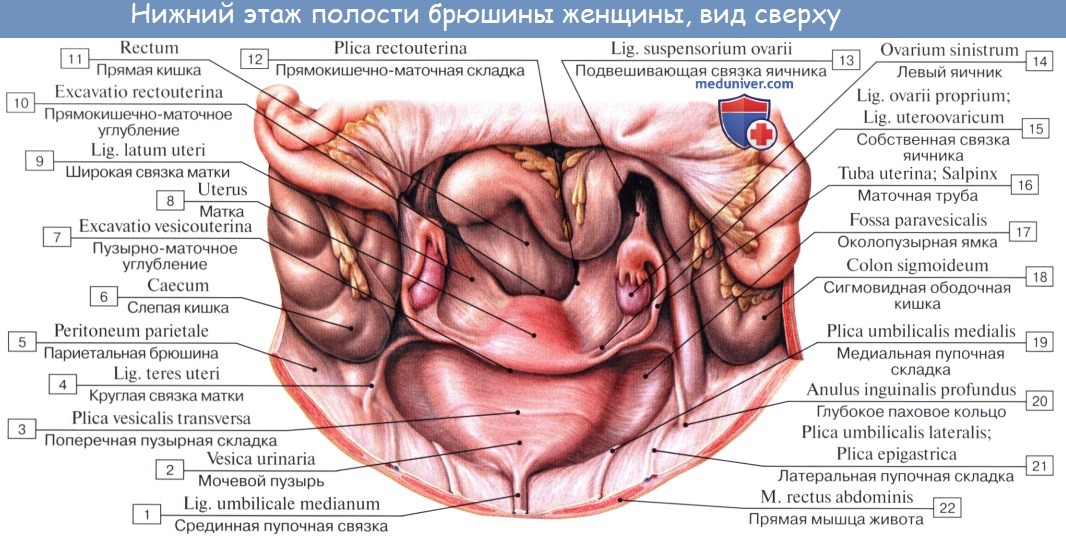
Физиологическое положение матки, труб и яичников обеспечивается подвешивающим, фиксирующим и поддерживающим аппаратами, Объединяющими брюшину, связки и тазовую клетчатку. Подвешивающий аппарат представлен парными образованиями, в него входят круглые и широкие связки матки, собственные связки и подвешивающие связки яичников. Широкие связки матки, собственные и подвешивающие связки яичников удерживают матку в срединном положении. Круглые связки притягивают дно матки кпереди и обеспечивают ее физиологический наклон.
Фиксирующий (закрепляющий) аппарат обеспечивает положение шатки в центре малого таза и делает практически невозможным ее смещение в стороны, кзади и кпереди. Но поскольку связочный аппарат отходит от матки в ее нижнем отделе, возможны наклонения матки в различные стороны. К фиксирующему аппарату относятся связки, расположенные в рыхлой клетчатке таза и идущие от нижнего отдела матки к боковым, передней и задней стенкам таза: крестцово-магочные, кардинальные, маточно-пузырные и пузырно-лобковые связки.
Кроме mesovarium, различают следующие связки яичников:
- подвешивающая связка яичника, раньше обозначавшаяся как воронкотазовая. Она представляет собой складку брюшины с проходящими в ней кровеносными (a. et v. ovarica) и лимфатическими сосудами и нервами яичника, натянутую между боковой стенкой таза, поясничной фасцией (в области деления общей подвздошной артерии на наружную и внутреннюю) и верхним (трубным) концом яичника;
- собственная связка яичника проходит между листками широкой маточной связки, ближе к заднему листку, и соединяет нижний конец яичника с боковым краем матки. К матке собственная связка яичника прикрепляется между началом маточной трубы и круглой связки, кзади и кверху от последней. В толще связки проходят rr. ovarii, являющиеся концевыми ветвями маточной артерии;
- аппендикулярно-яичниковая связка Кладо тянется о г брыжейки червеобразного отростка к правому яичнику или широкой связке матки в виде складки брюшины. Связка непостоянна и наблюдается у 1/2 – 1/3 женщин.
Поддерживающий аппарат представлен мышцами и фасциями тазового дна, разделенными на нижний, средний и верхний (внутренний) слои.
Наиболее мощным является верхний (внутренний) мышечный слой, представленный парной мышцей, поднимающей задний проход. Она состоит из мышечных пучков, которые расходятся веером от копчика к костям таза в трех направлениях (лобково-копчиковая, подвздошно-копчиковая и седалищно-копчиковая мышцы). Этот слой мышц называют также диафрагмой таза.
Средний слой мышц располагается между симфизом, лобковыми и седалищными костями. Средний слой мышц – мочеполовая диафрагма – занимает переднюю половину выхода таза, через него проходит мочеиспускательный канал и влагалище. В переднем отделе между ее листками находятся мышечные пучки, образующие наружный сфинктер мочеиспускательного канала, в заднем отделе заложены мышечные пучки, идущие в поперечном направлении, – глубокая поперечная мышца промежности.
Нижний (наружный) слой мышц тазового дна состоит из поверхностных мышц, форма расположения которых напоминает цифру 8. К ним относятся луковично-пещеристая, седалищно-кавернозная, наружный сфинктер заднего прохода, поверхностная поперечная мышца промежности.
Онтогенез яичников
Процесс роста и атрезии фолликулов начинается с 20 недель беременности, и к моменту родов в яичниках девочки остается до 2 млн ооцитов. К моменту менархе их количество уменьшается до 300 тыс. За весь период репродуктивной жизни достигает зрелости и овулирует не более 500 фолликулов. Начальный рост фолликулов не зависит от стимуляции ФСГ, ограничен, и быстро наступает атрезия. Полагают, что вместо стероидных гормонов основным регулятором роста и атрезии первичных фолликулов являются местные аутокринно/паракринные пептиды. Полагают, что процесс роста и атрезии фолликулов не прерывается никакими физиологическими процессами. Этот процесс продолжается во всех возрастах, включая внутриутробный период и менопаузу, непрерывается беременностью, овуляцией и ановуляцией. Механизм, который запускает рост фолликулов и их количество в каждом конкретном цикле, пока не ясен.
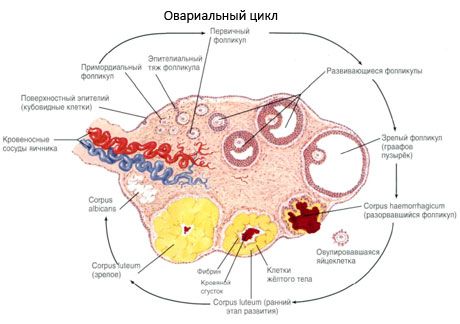
В своем развитии фолликул претерпевает несколько стадий развития. Примордиальные зародышевые клетки происходят из эндодермы желточного мешка, аллантоиса и мигрируют в генитальную область эмбриона на 5-6 неделе беременности. В результате быстрого митотического деления, которое продолжается от 6-8недель до 16-20недель беременности, в яичниках эмбриона образуется до 6-7 млн ооцитов, окруженных тонким слоем гранулезных клеток.
Преантральный фолликул – ооцит окружен мембраной (Zona pellucida). Гранулезные клетки, окружающие ооцит, начинают пролиферировать, их рост зависит от гонадотропинов и коррелирует с уровнем эстрогенов. Гранулезные клетки являются мишенью Для ФСГ. На стадии преантрального фолликула гранулезные клетки способны синтезировать три класса стероидов: преимущественно индуцирует активность ароматазы, основного фермента, превращающего андрогены в эстрадиол. Полагают, что эстрадиол способен увеличивать число собственных рецепторов, оказывая прямой митогенный эффект на гранулезные клетки независимый от ФСГ. Его рассматривают как паракринный фактор, усиливающий эффекты ФСГ, включая активизацию процессов ароматизации.
Рецепторы ФСГ появляются на мембранах гранулезных клеток сразу же как начинается рост фолликула. Снижение или увеличение ФСГ ведет к изменению числа его рецепторов. Это действие ФСГ модулируется ростовыми факторами. ФСГ действует через G-протеин, аденилат-циклазную систему хотя стероидогенез в фолликуле в основном регулируется ФСГ, в этот процесс вовлечены многие факторы: ионные каналы, рецепторы тирозин-киназы фосфолипазная система вторичных мессенжеров.
Роль андрогенов в раннем развитии фолликула сложна. На гранулезных клетках есть рецепторы андрогенов. Они являются не только субстратом для ФСГ-вызванной ароматизации в эстрогены, но могут в низких концентрациях усиливать процесс ароматизации. Когда уровень андрогенов увеличивается, преантральные гранулезные клетки преимущественно выбирают не путь ароматизации в эстрогены а более простой путь превращения в андрогены через 5а-редуктазу превращаясь в андроген, который не может быть превращен в эстроген, и таким путем ингибируется ароматазная активность. Этот процесс также ингибирует ФСГ и образование рецепторов ЛГ, таким образом, останавливая развитие фолликула.
Процесс ароматизации, фолликул с высоким уровнем андрогенов подвергается процессам атрезии. Рост и развитие фолликула зависит от его способности превращать андрогены в эстрогены.
В присутствии ФСГ доминантной субстанцией фолликулярной жидкости будут эстрогены. При отсутствии ФСГ – андрогены. ЛГ в норме в фолликулярной жидкости отсутствует до середины цикла. Как только увеличивается уровень ЛГ митотическая активность гранулезных клеток снижается, появляются дегенеративные изменения и увеличивается уровень андрогенов в фолликуле Уровень стероидов в фолликулярной жидкости выше, чем в плазме и отражает функциональную активность клеток яичников: гранулезных и тека-клеток. Если единственной мишенью для ФСГ являются гранулезные клетки, то у ЛГ имеется много мишеней – это тека-клетки, стромальные и лютеиновые клетки и гранулезные клетки. Способностью к стероидогенезу обладают и гранулезные и тека-клетки, но ароматазная активность преобладает в клетках гранулезы.
В ответ на ЛГ тека-клетки продуцируют андрогены, которые затем, через ФСГ – вызванную ароматизацию, превращаются гранулезными клетками в эстрогены.
По мере роста фолликула тека-клетки начинают экспрессировать гены для ЛГ-рецепторов, Р450 sec и 3бета-гидроксистероид-дегидрогеназу, инсулиноподобный фактор роста (IGF-1) синергично с ЛГ увеличивает экспрессию гена, однако не стимулируют стероидогенез.
Яичниковый стероидогенез всегда ЛГ-зависим. По мере роста фолликула тека-клетки экспрессируют Р450с17-энзим, который образует из холестерола андроген. Гранулезные клетки не имеют этого энзима и зависимы от тека-клеток в продукции эстрогенов из андрогенов. В отличие от стероидогенеза – фолликулогенез зависит от ФСГ. По мере роста фолликула и увеличения уровня эстрогенов приходит в действие механизм обратной связи – тормозится продукция ФСГ, что, в свою очередь, ведет к снижению ароматазной активности фолликула и, в конечном результате, к атрезии фолликула через апоптоз (программированную гибель клетки).
Механизм обратной связи эстрогенов и ФСГ ингибирует развитие начавших рост фолликулов, но не доминантного фолликула. Доминантный фолликул содержит больше рецепторов к ФСГ, которые поддерживают пролиферацию гранулезных клеток и ароматизацию андрогенов в эстрогены. Кроме этого действует паракринный и аутокринный путь, как важный координатор развития антрального фолликула.
Составной частью аутокринно/паракринного регулятора являются пептиды (ингибин, активин, фоллистатин), которые синтезируются гранулезными клетками в ответ на действие ФСГ и поступают в фолликулярную жидкость. Ингибин снижает ФСГ секрецию; активин стимулирует высвобождение ФСГ из гипофиза и усиливает действие ФСГ в яичнике; фоллистатин подавляет ФСГ активность, возможно за счет связывания активина. После овуляции и развития желтого тела ингибин находится под контролем ЛГ.
Рост и дифференциация овариальных клеток находится под влиянием инсулино-подобных факторов роста (IGE). IGF-1 воздействует на гранулезные клетки, вызывая увеличение циклического аденозин-монофосфата (цАМФ), прогестерона, окситоцина, протеогликана и ингибина.
IGF-1 действует на тека-клетки, вызывая увеличение продукции андрогенов. Тека-клетки, в свою очередь, продуцируют фактор некроза опухоли (tumor necrosis factor TNF) и эпидермальный фактор роста (EGF), которые также регулируются ФСГ.
EGF стимулирует пролиферацию гранулезных клеток. IGF-2 – основной фактор роста фолликулярной жидкости, в ней также обнаружены IGF-1, TNF-a, TNF-3 и EGF.
Нарушение паракринного и/или аутокринного регулирования овариальной функции, по-видимому, играет роль в нарушениях процессов овуляции и в формировании поликистозных яичников.
По мере роста антрального фолликула увеличивается содержание эстрогенов в фолликулярной жидкости. На пике их увеличения на гранулезных клетках появляются рецепторы к ЛГ, происходит лютеинизация гранулезных клеток и усиливается продукция прогестерона. Таким образом, в преовуляторный период увеличение продукции эстрогенов вызывает появление рецепторов ЛГ, ЛГ, в свою очередь, вызывает лютеинизацию гранулезных клеток и продукцию прогестерона. Увеличение прогестерона снижает уровень эстрогенов, что, по-видимому, вызывает второй пик ФСГ в середине цикла.
Полагают, что овуляция наступает через 10-12 часов после пика ЛГ и 24-36 часов после пика эстрадиола. Считается, что ЛГ стимулирует редукционное деление ооцита, лютеинизацию гранулезных клеток, синтез прогестерона и простагландина в фолликуле.
Прогестерон усиливает активность протеолитических энзимов, вместе с простагландином участвующих в разрыве стенки фолликула. Прогестероном вызванный пик ФСГ, позволяет выходу ооцита из фолликула путем превращения плазминогена в протеолитический энзим – плазмин, обеспечивает достаточное количество рецепторов ЛГ для нормального развития лютеиновой фазы.
В течение 3-х дней после овуляции гранулезные клетки увеличиваются, в них появляются характерные вакуоли, наполненные пигментом, – лютеином. Тека-лютеиновые клетки дифференцируются из теки и стромы и становятся частью желтого тела. Очень быстро под влиянием факторов ангиогенеза идет развитие капилляров, пронизывающих желтое тело, а с улучшением васкуляризации увеличивается продукция прогестерона и эстрогенов. Активность стероидогенеза и длительность жизни желтого тела определяется уровнем ЛГ. Желтое тело не является гомогенным клеточным образованием. Помимо 2-х типов лютеальных клеток в нем есть эндотелиальные клетки, макрофаги, фибробласты и др. Большие лютеальные клетки продуцируют пептиды (релаксин, окситоцин) и более активны в стероидогенезе с большей ароматазной активностью и большим синтезом прогестерона, чем малые клетки.
Пик прогестерона наблюдается на 8-й день после пика ЛГ. Отмечено, что прогестерон и эстрадиол в лютеиновую фазу секретируются эпизодически в корреляции с пульсовым выходом ЛГ. С образованием желтого тела контроль над продукцией ингибина переходит от ФСГ к ЛГ. Ингибин увеличивается вместе с увеличением эстрадиола до пика ЛГ и продолжает увеличиваться после пика ЛГ, хотя уровень эстрогенов снижается. Хотя ингибин и эстрадиол секретируются гранулезными клетками, они регулируются разными путями. Снижение ингибина в конце лютеиновой фазы вносит свой вклад в увеличение ФСГ для следующего цикла.
Желтое тело очень быстро – на 9-11 день после овуляции уменьшается.
Механизм дегенерации не ясен и не связан с лютеолитической ролью эстрогенов или с рецепторно-связанным механизмом, как это наблюдается в эндометрии. Есть другое объяснение роли эстрогенов, продуцируемых желтым телом. Известно, что для синтеза рецепторов прогестерона в эндометрии требуются эстрогены. Эстрогены лютеиновой фазы возможно необходимы для прогестерон-связанных изменений в эндометрии после овуляции. Неадекватное развитие рецепторов прогестерона, как следствие неадекватного содержания эстрогенов, является, возможно, дополнительным механизмом бесплодия и ранних потерь беременности, другой формой неполноценности лютеиновой фазы. Полагают, что длительность жизни желтого тела устанавливается в момент овуляции. И оно непременно будет подвергнуто регрессии, если не будет поддержано хорионического гонадотропина в связи с беременностью. Таким образом, регрессия желтого тела ведет к снижению уровней эстрадиола, прогестерона и ингибина. Снижение ингибина снимает его подавляющее влияние на ФСГ; снижение эстрадиола и прогестерона позволяет очень быстро восстановить секрецию ГнРГ и снять механизм обратной связи с гипофиза. Снижение ингибина и эстрадиола, совместно с увеличением ГнРГ дает превалирование ФСГ над ЛГ. Увеличение ФСГ приводит к росту фолликулов с последующим выбором доминантного фолликула, и начинается новый цикл, в том случае если не наступила беременность. Стероидные гормоны играют ведущую роль в репродуктивной биологии и в общей физиологии. Они определяют фенотип человека, влияют на сердечно-сосудистую систему, метаболизм костей, кожи, общее самочувствие организма и играют ключевую роль при беременности. Действие стероидных гормонов отражает внутриклеточные и генетические механизмы, которые необходимы для передачи внеклеточного сигнала к ядру клетки, чтобы вызвать физиологический ответ.
Эстрогены диффузно проникают через клеточную мембрану и связываются с рецепторами, расположенными в ядре клетки. Комплекс рецептор-стероид затем связывается с ДНК. В клетках мишенях эти взаимодействия приводят к экспрессии генов, синтезу протеинов, к определенной функции клеток и тканей.
Источник
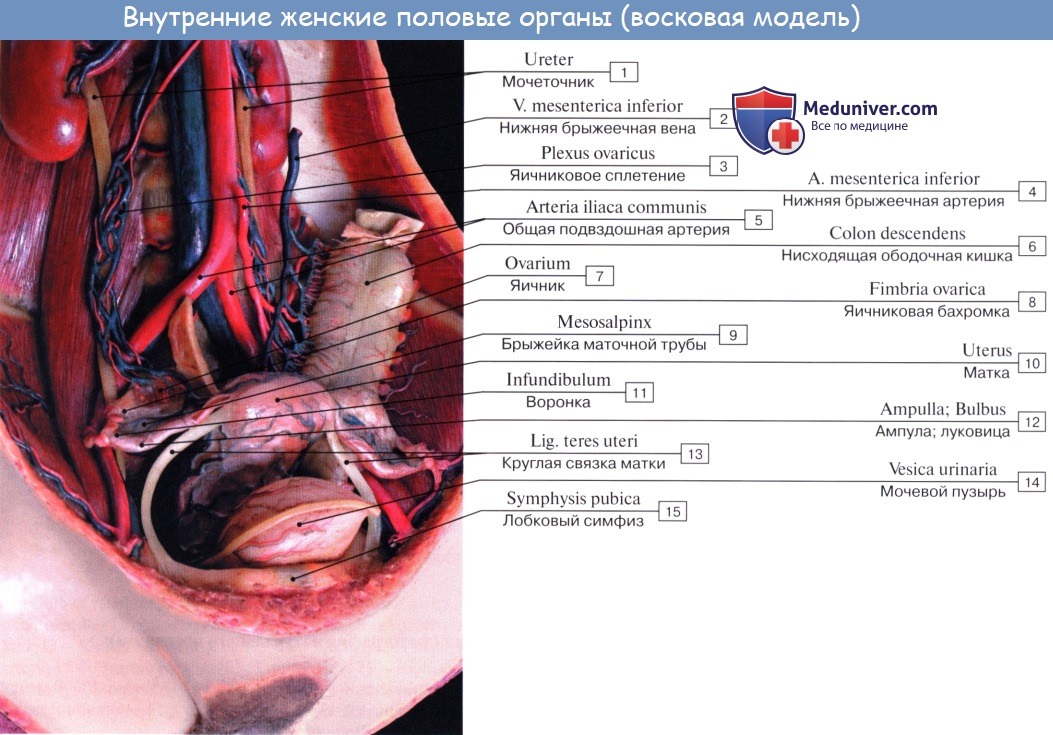
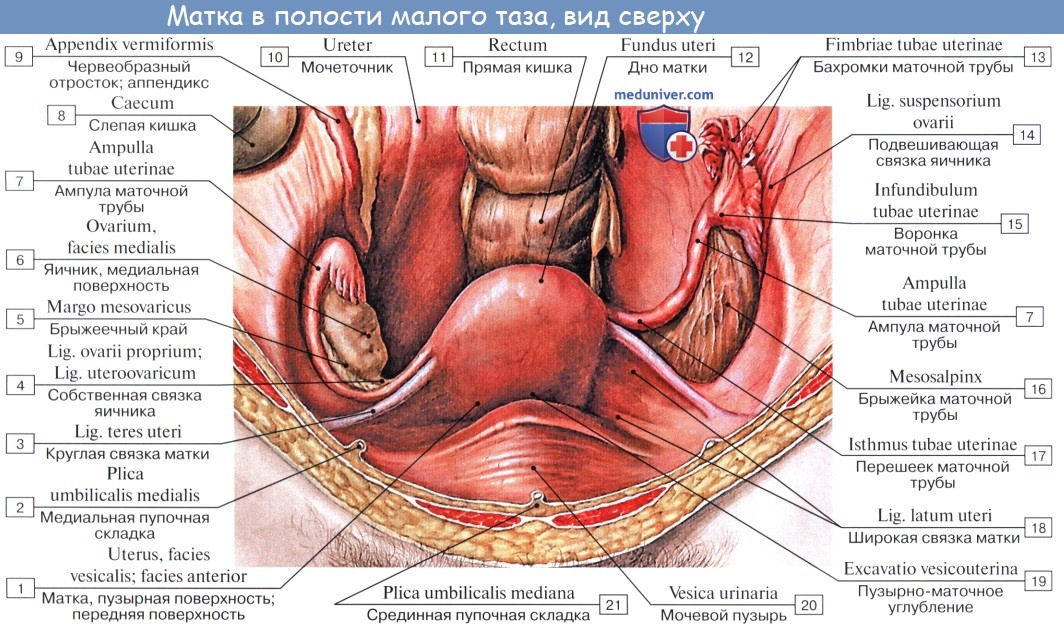
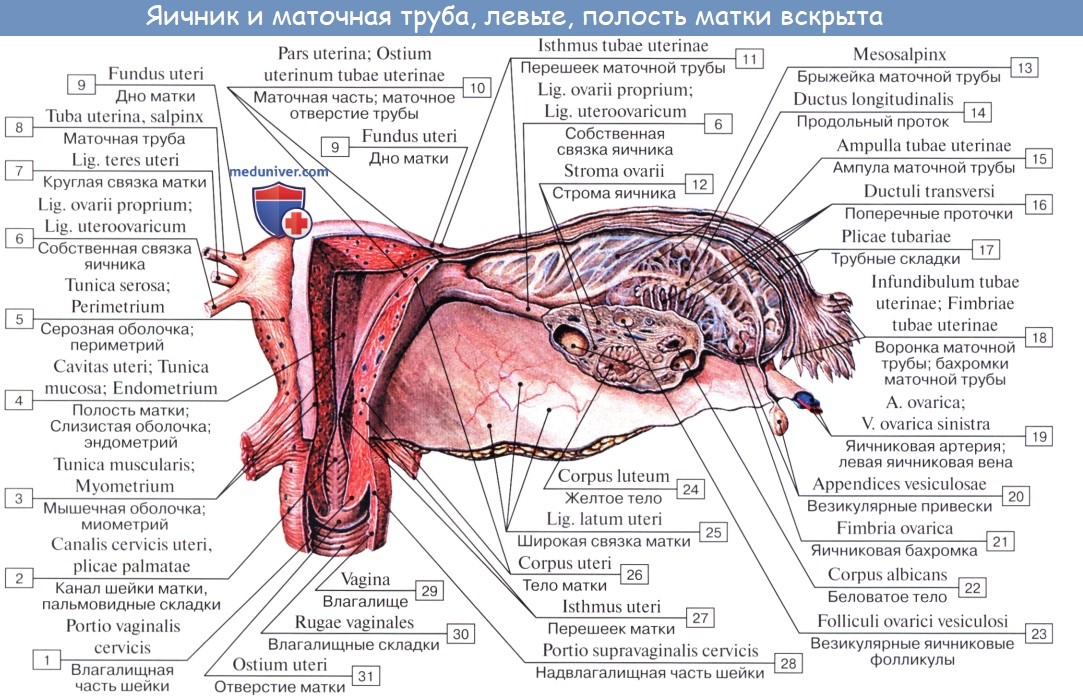
Оглавление темы “Женские половые органы.”: 1. Женские половые органы. Анатомия яичника. 2. Сосуды и нервы яичника. Кровоснабжение яичника. Иннервация яичника. 3. Маточная труба. Строение стенки трубы. Придаток яичника и околояичник. 4. Матка. Стенки матки. 5. Топография матки. Сосуды ( кровоснабжение ) матки. Иннервация ( нервы )матки. 6. Влагалище. Стенки влагалища. 7. Сосуды ( кровоснабжение ) влагалища. Нервы ( иннервация ) влагалища. 8. Наружные женские половые органы. Женская половая область. Лобок. Женские половые органы, organa genitalia femininaЖенские половые органы, organa genitalia feminina, состоят из двух отделов: 1) расположенные в тазу внутренние половые органы — яичники, маточные трубы, матка, влагалище и 2) видимый снаружи отдел — наружные половые органы (pudendum femininum), куда входят большие и малые половые губы, клитор, девственная плева.
Анатомия яичникаЯичник, ovarium , парный орган, является женской половой железой, аналогичной мужскому яичку. Он представляет плоское овальное тело длиной 2,5 см, шириной 1,5 см, толщиной 1 см. В нем различают два конца: верхний, несколько закругленный, конец обращен к маточной трубе и носит название трубного конца, extremitas tubaria; противоположный нижний, более заостренный, конец, extremitas uterina, соединен с маткой особой связкой (lig. ovarii proprium). Две поверхности, facjes lateralis et medialis, отделены друг от друга краями: свободный задний край, margo liber, выпуклый, другой же, передний край, брыжеечный, margo mesovaricus, прямой, прикрепляется к брыжейке. Этот край называют воротами яичника, hilus ovarii, так как здесь в яичник входят сосуды и нервы. Латеральной поверхностью яичник прилежит к боковой стенке таза между vasa iliaca externa и m. psoas major сверху, lig. umbilicale laterale спереди и мочеточником сзади. Длинник яичника расположен вертикально. Медиальная сторона обращена в сторону тазовой полости, но на значительном протяжении покрыта трубой, которая идет сперва вверх по брыжеечному краю яичника, затем на его трубном конце заворачивается и направляется вниз по свободному краю яичника. С маткой яичник связан посредством собственной связки, lig. ovarii proprium, которая представляет круглый тяж, заключенный между двумя листками широкой связки матки и состоящий в основном из непроизвольных мышечных волокон, продолжающихся в мускулатуру матки; собственная связка яичника тянется от маточного конца яичника к латеральному углу матки. Яичник имеет короткую брыжейку, mesovarium, представляющую собой дупликатуру брюшины, посредством которой он по своему переднему краю прикреплен к заднему листку широкой связки матки.
К верхнему трубному концу яичника прикрепляются fimbria ovarica (наиболее крупная из бахромок, окружающих брюшной конец трубы), а также треугольной формы складка брюшины — lig. suspensorium ovarii, спускающаяся к яичнику сверху от linea terminalis pelvis и заключающая яичниковые сосуды и нервы. Яичник содержит просвечивающиеся на свежем препарате везикулярные яичниковые фолликулы, folliculi ovarici vesiculosi, в каждом из которых находится развивающаяся женская половая клетка — ооцит. Фолликулы находятся в строме, stroma ovarii, в которой проходят сосуды и нервы. В зависимости от стадии развития фолликулы имеют различную величину — от микроскопических размеров до 6 мм в диаметре. Когда зрелый фолликул лопается (овуляция) и выделяется заключенный в нем ооцит, стенки его спадаются, полость выполняется кровью и клетками желтоватой окраски — получается желтое тело, corpus luteum. Ооцит превращается в зрелую яйцеклетку уже после овуляции в маточной трубе. В случае наступления беременности желтое тело увеличивается и превращается в крупное, около 1 см в диаметре, образование, corpus luteum graviditatis, следы которого могут сохраняться годами; желтое же тело, образующееся при отсутствии оплодотворения вышедшего из фолликула яйца, отличается меньшими размерами и через несколько недель исчезает. Вместе с атрофией клеток желтого тела последнее теряет свой желтый цвет и получает название белого тела, corpus albicans. С течением времени corpus albicans совершенно исчезает. Обыкновенно в течение 28 дней достигает зрелости один фолликул. Вследствие того, что фолликулы периодически лопаются (овуляция), поверхность яичника с возрастом покрывается морщинками и углублениями (о роли желтого тела см. «Эндокринные железы»). Яичник не покрыт брюшиной, которая здесь редуцировалась, а вместо нее он покрыт зародышевым эпителием. Благодаря этому яйцеклетка, после того как фолликул лопнул, может сразу попасть на поверхность яичника и далее в маточную трубу.
Учебное видео анатомии яичникаАнатомия яичника на препарате трупа от доцента Т.П. Хайруллиной разбирается Здесь – Также рекомендуем “Сосуды и нервы яичника. Кровоснабжение яичника. Иннервация яичника.” |

Источник