
Сосуды и ситовидные трубки находятся

Появление проводящих тканей в процессе эволюции является одной из причин, которые сделали возможным выход растений на сушу. В нашей статье мы рассмотрим особенности строения и функционирования ее элементов – ситовидных трубок и сосудов.
Особенности проводящей ткани
Когда на планете произошли серьезные изменения климатических условий, растениям пришлось приспосабливаться к ним. До этого все они обитали исключительно в воде. В наземно-воздушной среде стала необходимой добыча воды из почвы и ее транспортировка ко всем органам растения.
Различают два вида проводящей ткани, элементами которой являются сосуды и ситовидные трубки:
- Луб, или флоэма – расположена ближе к поверхности стебля. По ней органические вещества, образованные в листе во время фотосинтеза, передвигаются по направлению к корню.
- Второй тип проводящей ткани называется древесина, или ксилема. Она обеспечивает восходящий ток: от корня к листьям.

Ситовидные трубки растений
Это проводящие клетки луба. Между собой они разделены многочисленными перегородками. Внешне их строение напоминает сито. Отсюда и происходит название. Ситовидные трубки растений живые. Это объясняется слабым давлением нисходящего тока.
Их поперечные стенки пронизаны густой сетью отверстий. А клетки содержат много сквозных отверстий. Все они являются прокариотическими. Это означает, что в них нет оформленного ядра.
Живыми элементы цитоплазмы ситовидных трубок остаются только на определенное время. Продолжительность этого периода варьирует в широких пределах – от 2 до 15 лет. Данный показатель зависит от вида растения и условий его произрастания. Ситовидные трубки транспортируют воду и органические вещества, синтезированные в процессе фотосинтеза от листьев к корню.

Сосуды
В отличие от ситовидных трубок, эти элементы проводящей ткани представляют собой мертвые клетки. Визуально они напоминают трубочки. Сосуды имеют плотные оболочки. С внутренней стороны они образуют утолщения, которые имеют вид колец или спиралей.
Благодаря такому строению сосуды способны выполнять свою функцию. Она заключается в передвижении почвенных растворов минеральных веществ от корня к листьям.
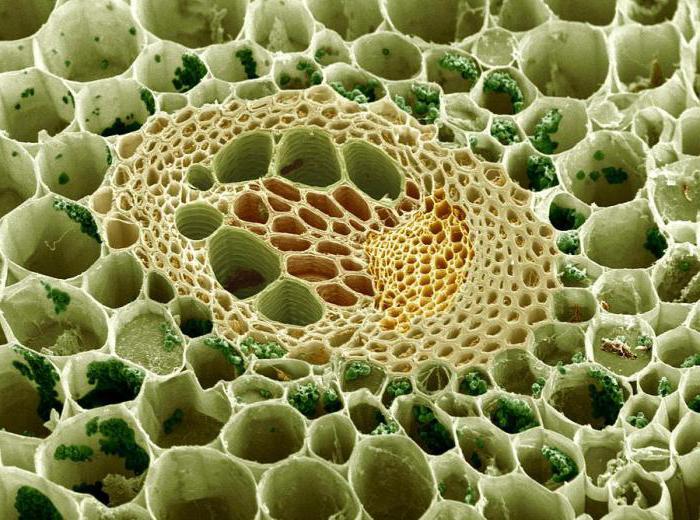
Механизм почвенного питания
Таким образом, в растении одновременно осуществляется передвижение веществ в противоположных направлениях. В ботанике этот процесс называют восходящим и нисходящим током.
Но какие силы заставляют воду из почвы двигаться вверх? Оказывается, что это происходит под влиянием корневого давления и транспирации – испарения воды с поверхности листьев.
Для растений этот процесс является жизненно необходимым. Дело в том, что только в почве находятся минералы, без которых развитие тканей и органов будет невозможным. Так, азот необходим для развития корневой системы. В воздухе этого элемента предостаточно – 75 %. Но растения не способны фиксировать атмосферный азот, поэтому минеральное питание так важно для них.
Поднимаясь, молекулы воды плотно сцепляются между собой и стенками сосудов. При этом возникают силы, способные поднять воду на приличную высоту – до 140 м. Такое давление заставляет почвенные растворы через корневые волоски проникать в кору, и далее к сосудам ксилемы. По ним вода поднимается к стеблю. Далее, под действием транспирации, вода поступает в листья.
В жилках рядом с сосудами находятся и ситовидные трубки. Эти элементы осуществляют нисходящий ток. Под воздействием солнечного света в хлоропластах листа синтезируется полисахарид глюкоза. Это органическое вещество растение расходует на осуществление роста и процессов жизнедеятельности.
Итак, проводящая ткань растения обеспечивает передвижение водных растворов органических и минеральных веществ по растению. Ее структурными элементами являются сосуды и ситовидные трубки.
Источник
«В природе нет ничего бесполезного» – Мишель де Монтень
Только вдумайтесь в мощь проводящей ткани! Ведь ей приходится поднимать воду и растворенные в ней минеральные вещества от тончайших волосков корня
до клеток листа. Самое высокое дерево на нашей планете, вечнозеленая секвойя по имени Гиперион, растет на севере Калифорнии и достигает (на 2017 год) – 117 метров в высоту.
И вода по проводящим тканям преодолевает 117 метров высоты у этого растения, от корней к листьям! Она передвигается по структурам проводящих тканей против
силы тяжести, и сегодня вы узнаете о секрете, который таит это уникальное явление.
Запомните, чтобы глубоко изучить любую науку, нужно восхищаться ей, уметь удивляться и проявлять любопытство в этой сфере. В ботанике это можно делать
самыми разными путями: вы можете посетить ботанический сад, или, к примеру, приобрести микроскоп и рассматривать ткани и органы растений, самостоятельно приготавливая
микропрепараты.
Это действительно важно, поэтому я останавливаюсь на этом. Сам я получаю и всегда призываю своих учеников получать искреннее удовольствие от погружения в науку.
Надеюсь, что и вы разделите эту радость новых интересных знаний, я приложу к этому все усилия. Итак, начнем изучать проводящие ткани.

Проводящие ткани можно сравнить с кровеносной системой человека, которая пронизывает весь наш организм, доставляя питательные вещества к клеткам и удаляя продукты
обмена веществ из них.
Как уже было сказано, эти ткани служат для передвижения по организму растения растворенных питательных веществ. Имеется два направления тока: от корней к листьям
(восходящий ток) и от листьев к корням (нисходящий ток).
Логическим путем можно угадать многие научные факты, даже не зная их. К примеру, чем представлен восходящий ток? Что поднимается от корней к листьям? Это конечно же
вода и растворенные в ней минеральные вещества, они движутся по сосудам и трахеидам проводящей ткани – ксилемы (древесины).
От листьев к корням спускаются органические вещества, образовавшиеся в результате фотосинтеза в листьях, они движутся по ситовидным трубкам проводящей ткани – флоэмы (луба).
Несмотря на то, что настоящие проводящие ткани впервые появились у папоротникообразных, но у мхов в наличии имеются водоносные клетки, благодаря которым они могут накапливать воду, которая в процентном соотношении может составить до 25% от их массы. По этой причине во время Первой мировой войны мох сфагнум использовали в качестве перевозочного материала. Кроме того, он обладает бактерицидными свойствами.
В состав и ксилемы, и флоэмы входят как живые, так и мертвые клетки. Однако отметим, что в ксилеме мертвые клетки преобладают.
Ксилема (древесина)
Обеспечивает восходящий ток (от корней к листьям) воды и растворенных в ней минеральных солей. В толще проводящей ткани находятся отнюдь не только те самые трахеиды и
сосуды, ее пронизывают многочисленные механические волокна – древесинные, обеспечивающие каркасность и прочность. В ксилеме содержатся также запасающие структуры,
представленные древесинной паренхимой, где накапливаются питательные вещества. Давайте разберемся из каких гистологических элементов состоит ксилема.
- Трахеиды
- Сосуды
- Древесинные волокна (либриформ)
- Паренхимные клетки (древесинная паренхима)
Эволюционно наиболее древние структуры. Представлены прозенхимными (вытянутые, с заостренными концами), мертвыми клетками. Через них осуществляется передвижение
и фильтрация растворов из нижележащей трахеиды в вышележащую. Их одревесневшая утолщенная клеточная стенка имеет разнообразные формы: пористую,
спиралевидную, кольчатую.
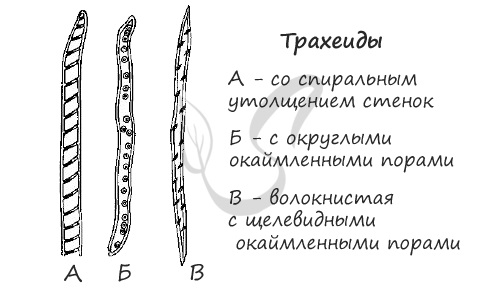
Длинные трубки, представляющие собой слияние отдельных мертвых клеток “члеников” в единый “сосуд”. Ток жидкости идет из нижележащих отделов в вышележащие
благодаря отверстиям (перфорациям) между клетками, составляющими сосуд. Так же, как и у трахеид, утолщения клеточных стенок у сосудов бывает самых разных форм.
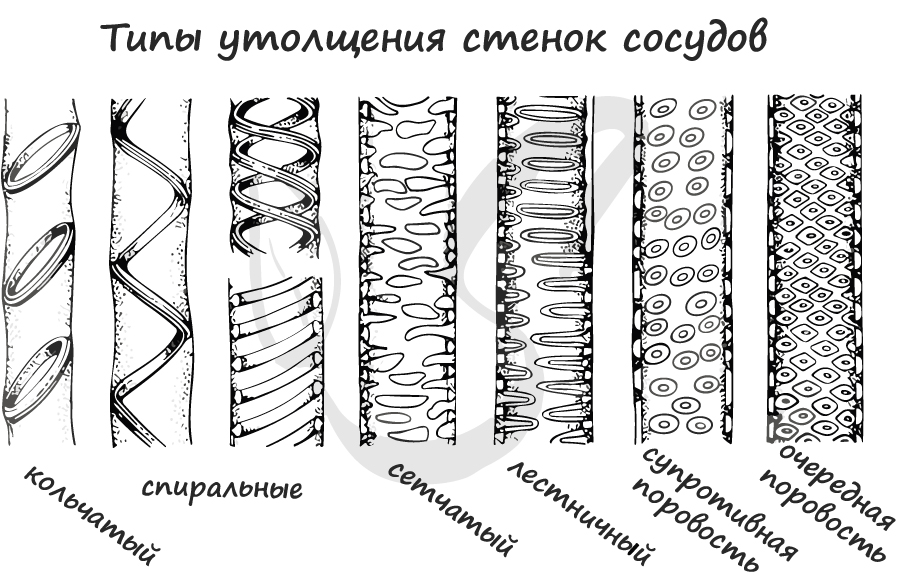
Во время роста растения проводящие ткани также претерпевают морфологические изменения. Изначальная длина сосуда меняется, благодаря своему строению
он растягивается и обеспечивает ток воды и минеральных солей.

Полагают, что эволюционно эти волокна берут начало от трахеид. Они не проводят воду, имеют более узкий просвет и отличаются хорошо выраженной
клеточной стенкой, которая придает ксилеме механическую прочность.
Эти клетки составляет обкладку вокруг сосуда, имеют одревесневшие оболочки с порами, которым соответствуют окаймленная пора со стороны сосуда. То есть сюда из сосуда могут поступать органические вещества и формировать запасы, которые в дальнейшем пригодятся растению.
Флоэма (луб)
Образовавшиеся в результате фотосинтеза в листьях продукты необходимо доставить в те части растения, где есть потребность в питательных веществах: конусы нарастания,
подземные части, или “складировать” на будущее в семенах и плодах. Флоэма обеспечивает нисходящий ток органических веществ в растении, доставляя их по месту назначения. До 90% всех перемещаемых веществ по флоэме составляет углевод – дисахарид сахароза.
Эта ткань представлена ситовидными трубками, генез (от греч. genesis – происхождение) которых различается: первичная флоэма дифференцируется из прокамбия, вторичная
флоэма – из камбия. Несмотря на различия генеза, клеточный состав описанных тканей идентичен.
Разберемся с компонентами, которые входят в состав флоэмы:
- Ситовидные элементы
- Склеренхимные элементы (лубяные волокна)
- Паренхимные элементы (лубяная паренхима)
Это живые клетки, обеспечивающие основной транспорт. Особо стоит выделить ситовидные трубки, образованные множеством безъядерных клеток – “члеников”, соединенных в единую цепь. Между “члениками” имеются поперечные перегородки с порами, благодаря которым содержимое из вышележащих клеток поступает в нижележащие. Эти перегородки похожи на сито – вот откуда берется название ситовидных трубок 🙂
Клетки-спутницы (сопровождающие клетки) также заслуживают нашего особого внимания. Они примыкают к боковым стенкам ситовидных трубок, из этих клеток через перфорации (поры) АТФ
и нуклеиновые кислоты попадают в ситовидные трубки, создавая нисходящий ток. Таким образом, клетки-спутницы контролируют деятельность
ситовидных трубок.
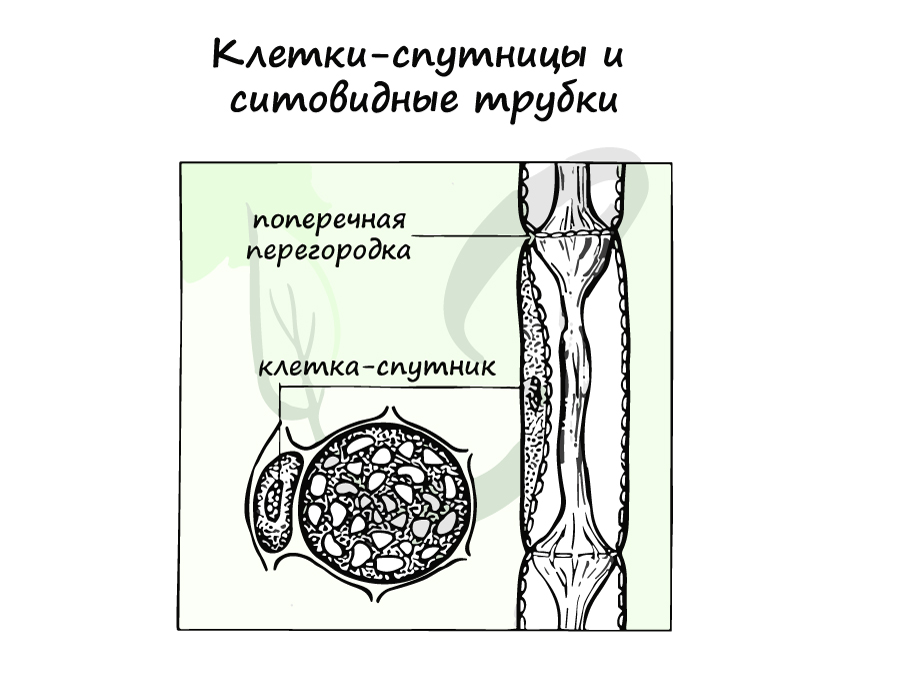
Пронизывают флоэму, придавая ей опору. Часть клеток отмирает, что характерно для данной группы тканей.
Обеспечивают радиальный транспорт веществ из проводящих тканей в рядом расположенные живые клетки других прилежащих тканей.
По мере старения ситовидные трубки закупориваются каллозой (образующей так называемое мозолистое тело) и затем отмирают.
Отмершие ситовидные трубки постепенно сплющиваются давящими на них соседними живыми клетками.
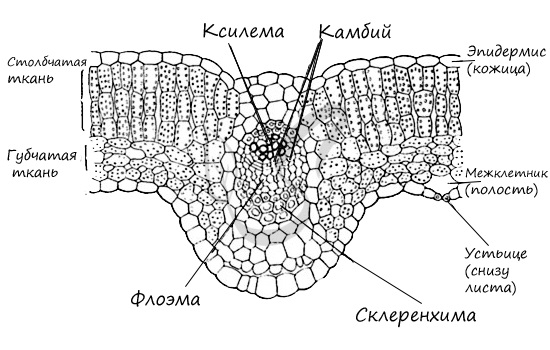
Ниже вы найдете продольный срез тканей растения, изучите его.
Жилка
Это сосудисто-волокнистый пучок, образованный ксилемой и флоэмой. Ксилема располагается сверху, флоэма – снизу. Над пучком и под ним располагаются уголковая или пластинчатая
колленхима, прилежащая к эпидерме и выполняющая опорную функцию. Склеренхима может располагаться участками или вокруг этих жилок. Жилки развиваются из прокамбия,
располагаются в центральном осевом цилиндре. Существует два вида жилок:
- Открытые
- Закрытые
Ключевой момент: между ксилемой и флоэмой располагается прослойка камбия. Этот факт обуславливает возможность образования дополнительного объема
ксилемы и флоэмы в будущем, для дальнейшего роста и увеличения в объеме пучка. Без камбия невозможно было бы утолщения органа. Такие пучки можно
обнаружить во всех органах двудольных растений.
Основное отличие в том, что между ксилемой и флоэмой отсутствует камбий. Невозможно образования новых элементов проводящих тканей, ксилемы и флоэмы.
Закрытые сосудисто-волокнистые пучки встречаются в стеблях однодольных растений.
Верхняя часть жилки представлена ксилемой, нижняя флоэмой. Вокруг пучка в виде кольца располагается механическая ткань
– склеренхима. Над пучком и под ним механическая ткань – колленхима – выполняет опорную функцию.
Как вода поднимается от корней к листьям, против силы тяжести?
Запомните, что вода и растворенные в ней минеральные соли поступают в растение благодаря слаженной работе двух концевых двигателей: нагнетающего корневого и
присасывающего листового.
- Корневое давление
- Транспирация
Силу, поднимающую воду вверх по сосудам, называют корневым давлением. Величина его обычно составляет от 30 до 150 кПа. В основе этого явления лежит осмос:
клетки корня выделяют минеральные и органические вещества в сосуды, что создает более высокое давление, чем в почвенном растворе, и последний начинает притягиваться
в сосуды.
Работа верхнего концевого двигателя заключается в транспирации – испарении воды с поверхности листа. Представим себе длинный сосуд с жидкостью от корневых
волосков до клеток листа. Далее проведите следующий мысленный эксперимент: из верхнего конца трубки жидкость все время удаляется путем испарения, то есть место
освобождается и это создает притягивающую силу для жидкости расположенной ниже, она поднимается наверх, на место испарившейся жидкости.
Присасывающее действие транспирации передается корням в форме гидродинамического натяжения, которое связывает между собой работу обоих двигателей.
© Беллевич Юрий Сергеевич 2018-2020
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Источник
Проводящая ткань — вид тканей растений, служащих для передвижения по организму растворённых питательных веществ. У многих высших растений она представлена проводящими элементами (сосудами и ситовидными трубками). В стенках проводящих элементов есть поры и сквозные отверстия, облегчающие передвижение веществ от клетки к клетке.
Проводящая ткань образует в теле растения непрерывную разветвлённую сеть, соединяющую все его органы в единую систему — от тончайших корешков до молодых побегов, почек и кончиков листа.
Происхождение[править | править код]
Учёные считают, что возникновение тканей связано в истории Земли с выходом растений на сушу. Когда часть растения оказалась в воздушной среде, а другая часть (корневая) — в почве, появилась необходимость доставки воды и минеральных солей от корней к листьям, а органических веществ от листьев к корням. Так в ходе эволюции растительного мира возникло два типа проводящих тканей — древесина и луб. По древесине (по трахеидам и сосудам) вода с растворёнными минеральными веществам поднимается от корней к листьям — это водопроводящий, или восходящий, ток. По лубу (по ситовидным трубкам) образовавшиеся в зелёных листьях органические вещества поступают к корням и другим органам растения — это нисходящий ток.
Значение[править | править код]
Проводящие ткани растений — это ксилема (древесина) и флоэма (луб). По ксилеме (из корня в стебель) идёт восходящий ток воды с растворёнными в ней минеральными солями. По флоэме — более слабый и медленный ток воды и органических веществ.
Значение древесины
Ксилема, по которой идёт сильный и быстрый восходящий ток, образована мёртвыми, разными по величине клетками. Цитоплазмы в них нет, стенки одревеснели и снабжены многочисленными порами. Представляют собой цепочки из прилегающих друг к другу длинных мёртвых водопроводящих клеток. В местах соприкосновения у них имеются поры, по которым и передвигаются растворы из клетки в клетку по направлению к листьям. Так устроены трахеиды.
У цветковых растений появляются и более совершенные проводящие ткани — сосуды. В сосудах поперечные стенки клеток в большей или меньшей степени разрушаются. Таким образом, сосуды — это полые трубки, образованные множеством мёртвых трубчатых клеток (члеников). По таким сосудам растворы передвигаются ещё быстрее. Помимо цветковых, другие высшие растения имеют только трахеиды.
Значение луба
В силу того, что нисходящий ток более слабый, клетки флоэмы могут оставаться живыми. Они образуют ситовидные трубки — их поперечные стенки густо пронизаны отверстиями. Ядер в таких клетках нет, но они сохраняют живую цитоплазму. Ситовидные трубки остаются живыми недолго, чаще 2-3 года, изредка — 10-15 лет. На смену им постоянно образуются новые ситовидные трубки.
Визуализация[править | править код]
Интересный метод визуализации проводящей системы деревьев предложили В. И. Иванов-Омский и Е. И. Иванова. Они использовали коронный разряд, или, точнее, эффект Кирлиана. Этим методом у осины, например, обнаружены эллипсовидные на срезе конгломераты сосудов[1].
См. также[править | править код]
- Концентрический пучок
- Коллатеральный пучок
- Биколлатеральный пучок
- Радиальный пучок
Примечания[править | править код]
- ↑ Иванов-Омский В.И., Иванова Е.И. Фотографирует разряд: древесный водопровод // Природа. — 2013. — № 3. — С. 14-19.
Источник
Ситовидныетрубки (решётчатыетрубки, ситовидныесосуды) —
проводящие
элементы
в
телах
высших
растений,
проводящие
сахар
и
пластические
питательные
вещества,
развитые
преимущественно
в
лубяной
части
сосудисто-волокнистого
пучка.
Главной
их
функцией
является
транспортировка
углеводов —
например,
из
листьев
в
плоды
и
корни.
Были
открыты
и
названы
германским
биологом
и
исследователем
леса Теодором
Гартигом в1837
году.
Ситовидные
трубки
представляют
однорядные
тяжи
вытянутых
в
длину
клеток,
толстые
и
различных
цветов;
их
конечные
стенки
являются
ситовидными
пластинками,
несущими
ситовидные
поля
с
многочисленными
прободениями,
которые
изнутри
выстланы
каллозой.
Простые
пластинки
обычно
горизонтальные
и
содержат
одно
ситовидное
поле
(характерными
примерами
являются
тыква
и
ясень),
сложные
же
расположены
под
наклоном
и
имеют
несколько
ситовидных
полей
(к
таким
растениям
относятся
липа,
виноград,
пассифлора,
рис).
К
каждому
членику
ситовидной
трубки
примыкает
тяж
узких
сопровождающих
клеток,
которые
физиологически
связаны
с
ними[1].
При
их
развитии
тонопласты
в
клетках
постепенно
разрушаются,
вызывая
смешивание
цитоплазмы
с
клеточным
соком;
происходит
дегенерация
органелл
и
ядра
клетки.
Ввиду
значительного
количества
цитоплазмы
не
считаются
«настоящими»
клетками,
они
не
имеют
клеточного
ядра,
рибосом
и
вакуоли.
У
большинства
растений
ситовидные
трубки
функционируют
не
более
года,
но
встречаются
и
исключения:
у
винограда
они
существуют
2 года,
у
липы —
в
течение
нескольких
лет,
тогда
как
у
некоторых
пальм —
несколько
десятков.
В
конце
вегетационного
периода
ситовидные
прободения
полностью
закупориваются
каллозой,
откладывающейся
также
на
обеих
сторонах
ситовидной
пластинки,
вследствие
чего
образуются
мозолистые
тела.
Более
не
функционирующие
ситовидные
трубки
и
окружающие
клетки
со
временем
деформируются
и
облитерируются.
КАЛЛОЗА,
каллёза,
полисахарид,
содержащийся
в
растениях
и
состоящий
из
остатков
молекулы
глюкозы,
соединенных
Р-1,3
связями
в
спиральную
цепочку.
Каллоза
нерастворима
в
воде,
хорошо
флюоресцирует
в
люминесцентном
микроскопе.
У
виноградного
растения
каллоза
образуется
как
в
вегетативных,
так
и
в
генеративных
органах.
Она
выстилает
канальцы
ситовидных
пластинок
флоэмы
осенью
1-го
года
их
образования,
весной,
гидролизуясь,
рассасывается.
Осенью
2-го
года
каллоза
вновь
закупоривает
канальцы,
а
на
3-й
год
выводит
ситовидные
трубки
из
строя.
На
стенках
клеток
паренхимы
пораженных
органов
каллоза
способствует
образованию
каллуса.
В
клетках
фиброзного
слоя
пыльника
откладывающаяся
каллоза
образует
утолщения.
В
оболочках
микро-
и
макроспороцитов
(и
соответственно
микро-
и
макроспор)
каллоза
играет
роль
изолятора,
а
также
предохраняет
от
обезвоживания
и
др.
неблагоприятных
условий,
что
имеет
важное
значение
при
спорогенезе.
В
оболочке
растущих
пыльцевых
трубок,
особенно
в
базальной
части,
откладывающаяся
каллоза
вызывает
утолщения,
часто
—
каллозные
пробки.
20) Проводящие пучки растений
Обособленные
тяжи
проводящей
системы,
состоящие
чаще
из ксилемы и флоэмы ,
называют
проводящими
пучками.
Первоначально
они
возникают
из прокамбия .
Вокруг
пучков
нередко
формируется
обкладка
из
живых
или
мертвых паренхимных клеток.
Они
могут
быть
полными,
т.е.
состоящими
из флоэмы и ксилемы ,
и
неполными,
состоящими
только
из
ксилемы
или
флоэмы.
В
тех
случаях,
когда
часть прокамбия сохраняется
и
превращается
затем
в камбий ,
а
пучок
способен
к
вторичному
утолщению,
говорят
об
открытых
пучках.
Они
встречаются
у
большинства двудольных и голосеменных .
В
закрытых
пучках
однодольных
прокамбий
полностью
дифференцируется
в
проводящие
ткани.
В
зависимости
от
взаимного
расположения флоэмы и ксилемы различают
пучки
нескольких
типов.
Чаще
всего
флоэма
лежит
по
одну
сторону
от
ксилемы.
Такие
пучки
называют
коллатеральными
(открытые
и
закрытые).
У
части двудольных
растений(из
семейств пасленовых , вьюнковых , тыквенных и
т
д.)
одна
часть
флоэмы
располагается
снаружи,
а
другая
– с
внутренней
стороны
ксилемы.
Такой
пучок
называется
биколлатеральным,
a соответствующие
участки флоэмы –
наружной
и
внутренней
флоэмой. Камбий находится
между
наружной флоэмой и ксилемой .
Встречаются
также
концентрические
пучки,
при
этом
либо
флоэма
окружает
ксилему
(центроксилемные
пучки),
либо,
наоборот,
ксилема
окружает
флоэму
(центрофлоэмные).
Центрофлоэмные
пучки
найдены
в
стеблях
и
корневищах
ряда двудольных ( ревень ,щавель , бегония )
и однодольных (многие лилейные , осоковые ).
Известны
пучки
промежуточные
между
закрытыми
коллатеральными
и
центрофлоэмными.
Центроксилемные
пучки
обычны
для папоротников .
В
центре
молодых
корней голосеменных и покрытосеменных ,
имеющих
первичное
строение,
располагается
проводящий
пучок,
получивший
название
радиального. Ксилема в
таком
пучке
расходится
лучами
от
центра,
а флоэма располагается
между
лучами.
Возникают
эти
пучки
из прокамбия .
В
корнях двудольных и голосеменных между
ксилемой
и
флоэмой
сохраняется
слой
прокамбиальных
клеток,
которые
позднее
дифференцируются
в камбий .
Встречаются
однолучевые
(монархные),
двулучевые
(диархные),
трехлучевые
(триархные),
четырехлучевые
(тетрархные),
пятилучевые
(пентархные)
и
многолучевые
(полиархные)
радиальные
лучи.
Последние
обычны
у однодольных .
Ксилема
и
флоэма
обычно
расположены
рядом,
образуя
слои,
или
так
называемые проводящиепучки,
представленные
в
растениях
несколькими
типами.
В
зависимости
от
расположения
ксилемы
и
флоэмы
относительно
друг
друга,
различают
следующие
типы:
1. Коллатеральные (бокобочные),
когда
ксилема
и
флоэма
располагаются
бок
о
бок,
т.е.
на
одном
радиусе
(рис.
19, А,
В).
2. Биколлатеральные (дважды
бокобочные
пучки)
– флоэма
прилегает
к
ксилеме
с
обеих
сторон.
Наружный
участок
флоэмы
более
мощный
(рис.
19, Б).
3. Концентрические :
а) амфивазальные –
ксилема
замкнутым
кольцом
окружает
флоэму
(рис.
19, Г
);
б) амфикрибральные –
флоэма
окружает
ксилему
(рис.
19, Д).
4. Радиальные –
ксилема
расходится
лучами
от
центра,
а
флоэма
располагается
между
лучами
(рис.
19, Е).
Проводящие
пучки
в
зависимости
по
наличию
или
отсутствию
в
них
камбия
бывают открытые и закрытые.
В
открытых
– между
ксилемой
и
флоэмой
есть
камбий
(рис.
19, А,
Б).
В
закрытых –
камбия
нет
(рис.
19, В).
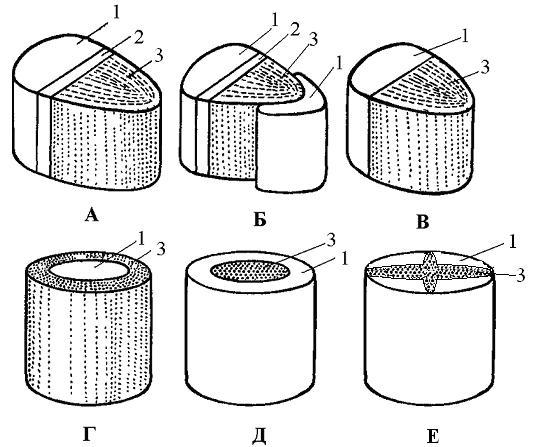
Рис.
19. Типы
проводящих
пучков:
А
– открытый
коллатеральный;
Б
– открытый
биколлатеральный;
В
– закрытый
коллатеральный;
Г,
Д
– концентрические
(Г
– амфивазальный,
Д
– амфикрибральный);
Е
– радиальный.
1
– флоэма,
2 – камбий,
3 – ксилема.
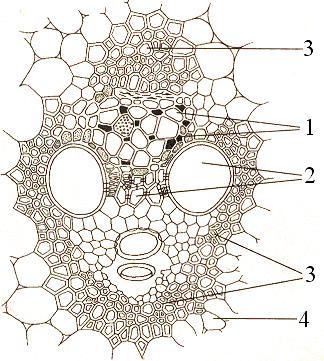
Рис.
20. Закрытый
коллатеральный
проводящий
пучок
на
поперечном
срезе
стебля
кукурузы:
1
– флоэма,
2 – ксилема,
3 – механическая
обкладка
пучка,
4 – основная
паренхима
стебля,
окружающая
пучок.
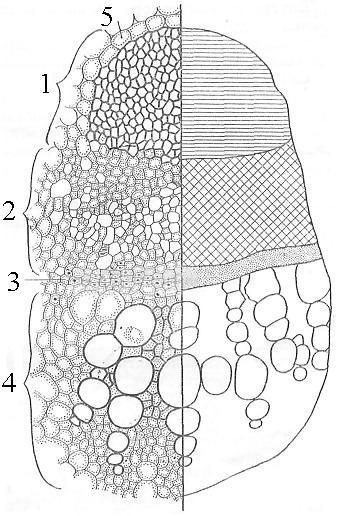
Рис.
21. Открытый
коллатеральный
проводящий
пучок
на
поперечном
срезе
стебля
подсолнечника
(слева
– детальный
рисунок,
справа
– схематичный):
1
– склеренхима,
2 – флоэма,
3 – камбий,
4 – ксилема,
5 – основная
паренхима
стебля.
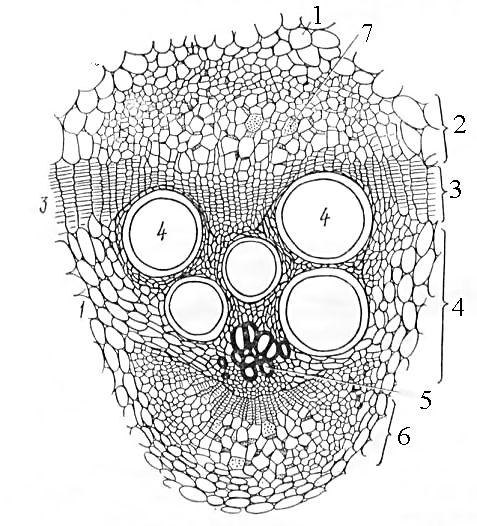
Рис.
22. Открытый
биколлатеральный
проводящий
пучок
на
поперечном
срезе
стебля
тыквы:
1
– основная
паренхима
стебля,
2 – наружная
флоэма,
3 – камбий,
4 – вторичная
ксилема,
5 – первичная
ксилема,
6 – внутренняя
флоэма,
7 – ситовидная
пластинка.
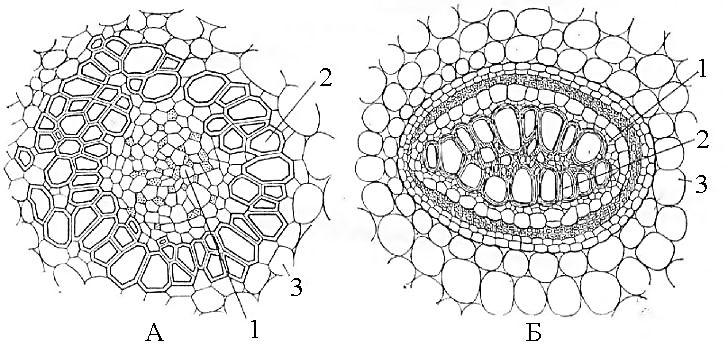
Рис.
23. Концентрические
проводящие
пучки:
А
– амфивазальный
пучок
корневища
ландыша;
Б
– амфикрибральный
пучок
корневища
папоротника-орляка.
1
– флоэма,
2 – ксилема,
3 – основная
паренхима
стебля.
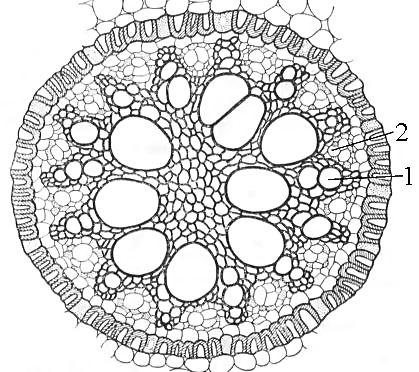
Рис.
24. Радиальный
проводящий
пучок
корня
ириса:
1
– луч
ксилемы,
2 – участок
флоэмы.
21)
К
выделительным
(секреторным)
тканям
относятся
разного
рода
структурные
образования,
способные
активно
выделять
из
растения
или
изолировать
в
его
тканях
продукты
метаболизма
и
капельно-жидкую
воду.
Выделяемые
наружу
или
накапливаемые
внутри
жидкие
и
твердые
продукты
метаболизма
получили
общее
название секретов .
Как
правило, секреты (смесь терпеноидов ,полифенольные
соединения , оксалат
кальция )
относятся
к
продуктам вторичного
метаболизма (обмена),
но
среди
них
встречаются
и
продукты первичного
обмена .
Вторичные
метаболиты представлены
в
растениях
огромным
числом
индивидуальных
соединений,
хотя
они
образуются
на
немногих
путях
обмена
веществ
( схема
1 )
и
их
биогенетическими
предшественниками
являются
– мевалонат , ацетил-КоA
(ацетил-коэнзим
A) –
сложное
органическое
вещество,
молекулы
которого
участвуют
в
главнейших
биохимических
реакциях,
идущих
в
живой
клетке,
коричная
кислота
и
ряд
белковых
аминокислот.
В
левой
части схемы
1 указаны
биогенетические
предшественники
основных
классов
соединений,
относимых
к
продуктамвторичного
метаболизма .
Три
предшественника: мевалонат , ацетил-КоA и аминокислоты относятся
к
продуктам
первичного обмена ; коричная
кислота –
продукт
уже
вторичного
обмена,
но
она
оказывается
необходимой
в
ходе
биохимического
синтеза флавоноидов и
различных
более
простых
природных
производных
фенилпропана,
обобщенно
называемых
фенилпропаноидами.
В
правой
колонке
схемы
перечислены
главнейшие
классы вторичных
метаболитов (цифрами
указано
приблизительное
число
индивидуальных
соединений,
выделенное
из
различных
групп
организмов).
Стрелками
показана
связь
между
теми
или
иными
соединениями
в
ходе
биохимических
реакций.
Сами
биохимические
реакции,
в
ходе
которых
одни
соединения
превращаются
в
другие,
довольно
разнообразны
и
включают
множество
стадий.
Элементы,
или
комплексы,
выделительных
тканей
встречаются
во
всех
органах.
В
зависимости
от
того,
выделяют
они
вещества
наружу
или
выделенные
вещества
остаются
внутри
растения,
их
делят
на
две
группы:
ткани
внутренней
и
наружной
секреции.
Клетки
выделительных
тканей
по
форме
обычно паренхимные и
тонкостенные.
Они
долго
остаются
живыми,
выделяя секрет .Клетки-идиобласты по
мере
накопления
большого
количества
секрета
лишаются протопласта и
стенки
их
нередко
опробковевают.
Синтез
жидких
секретов
связывают
с
деятельностью
внутриклеточных
мембран
и комплекса
Гольджи .
Поскольку
продукты вторичного
метаболизма биологически
активны
и
могут
вызвать
повреждение цитоплазмы ,
существуют
механизмы,
препятствующие
этому.
Один
из
них
– перенос
таких
веществ
в вакуоль или
в
свободное
изолированное
от
цитоплазмы
пространство
клетки.
Другой
механизм
– химическое
превращение
соединений
до
относительно
безвредных,
что,
разумеется,
не
исключает
их
последующее
выделение.
Прежде
чем
выделиться
из цитоплазмы ,
где
они
синтезируются, секретируемые
вещества преодолевают
цитоплазматические
мембраны
– плазмалемму ,
если
вещества
выделяются
в
свободное
пространство
клетки,
или тонопласт –
при
транспорте
в вакуоль .
Эволюционно
внутренние
выделительные
ткани
возникли
из ассимиляционных и запасающих ,
а
наружные
связаны
с покровными
тканями .
Клетки,
содержащие оксалат
кальция ,
изначально
выступают
как ассимиляционные или запасающие и
лишь
позднее
превращаются
в
выделительные.
Функции
выделительных
тканей
растений
существенно
отличаются
от
функций
выделительной
системы
животных.
Образующиеся
секреты
нередко
эффективно
защищают
растения
от
поедания
животными,
повреждения
насекомыми
или
патогенными
микроорганизмами.
Часто секреты ,
выступающие
из
мест
поранения
растений
при
искусственных
или
естественных
повреждениях,
играют
роль
бактерицидного
пластыря
(смолы,
бальзамы).
Выделяющиеся
в цветках ароматические
и
сахаристые
вещества
( нектар )
привлекают
насекомых-опылителей.
Наконец,
накапливающиеся
в
разного
рода
вместилищах
секретированные
вещества
могут
вновь
вовлекаться
в
процесс метаболизма и
в
этом
случае
выступают
в
роли
запасных
веществ. Клетки-идиобласты ,
особенно
содержащиеоксалат
кальция ,
приобретают
значение
мест
длительного
“захоронения”
токсичных
для
растения
веществ
или
веществ,
полностью
исключенных
из
метаболизма.
Вещества,
полностью
исключающиеся
из
метаболизма,
удаляются
из
растения
при
опадении
листьев,
слущивании
корки
и
т.п.
Это
основной
путь
избавления
от
“шлаков”.
Судьба
секретирующих
клеток
различна.
Иногда
они
остаются
живыми
длительное
время.
При
этом
секреция
осуществляется
путем
пассивного
или
активного
транспорта
либо экзоцитоза .
В
иных
случаях
при
секреции
происходит
повреждение
клетки.
Выделение
наружу
выработанного секрета сопровождается
выбросом
части цитоплазмы ,
но
отделяется
только
безъядерная
часть
клетки.
Наконец,
известны
случаи,
когда
клетка
полностью
дегенерирует
и
иногда
вместе
с
выработанным
ею
продуктом
выделяется
в
окружающую
среду
(например,
солевые
волоски
некоторых галофитов и
слизистые
клетки корневого
чехлика ).
Выделение
капельно-жидкой
воды
характерно
для
многих
растений
и
осуществляется
через гидатоды .
Ткани
внутренней
секреции
могут
быть
представлены
отдельными клетками-идиобластами , вместилищами
выделений , смоляными
ходами , эфирномасляными
каналами и млечниками ( рис.
49 ).
24)
Ка́мбий (от лат. cambium —
обмен,
смена),
образовательная
ткань
в
стеблях
и корнях преимущественно
двудольных
и
голосеменныхрастений,
дающая
начало
вторичным
проводящим
тканям
и
обеспечивающая
рост
их
в
толщину.
Сезонные
изменения
активности
камбия
обусловливают
образование
годичных
колец древесины.
образовательная
ткань
(Меристема)
в
корнях
и
стеблях
преимущественно
двудольных
и
голосеменных
растений.
К.
расположен
однорядным
цилиндрическим
слоем
(на
поперечном
срезе
—
в
виде
кольца).
В
результате
деятельности
К.
происходит
прирост
осевых
органов
в
толщину.
Он
образует
кнаружи
вторичную
флоэму
(См. Флоэма)
(луб)
и
кнутри
—
вторичную
ксилему
(См. Ксилема)
(древесину).
К.
возникает
из
клеток
прокамбия
(См. Прокамбий),
лежащих
между
первичными
флоэмой
и
ксилемой.
В
образовании
К.
в
корнях
большое
значение
имеет Перицикл.
В
листьях
К.
нет,
если
же
он
есть,
то
его
деятельность
рано
затухает.
По
форме
клетки
К.
прозенхимные
—
удлинённо-заострённые
(на
концах
скошены),
таблитчатые,
расположены
по
длине
органа
(рис.
1).
Оболочки
клеток
К.
мягкие,
целлюлозные,
имеют
первичные
поровые
поля
с
плазмодесмами
(См. Плазмодесмы)
Клетки
К.
делятся,
видоизменяясь
в
клетки
луба
(кнаружи
от
К.)
или
древесины
(кнутри
от
К.)
(рис.
2).
Обычно
древесины
образуется
в
несколько
раз
больше,
чем
луба.
В
результате
деления
некоторых
клеток
К.
образуются
мелкие
клетки:
это
—
лубо-древесинные
лучи
(рис.
3),
производящие
в
одну
сторону
лубяную,
в
др.—
древесинную
часть
луча.
На
осень
и
зиму
деятельность
К.
прекращается
(в
умеренных
широтах).
Периодичность
деятельности
К.
вызывает
образование
годичных
колец
(См. Годичные
кольца).
В
зависимости
от
характера
деления
клеток
К.
различают
ярусный
и
неярусный
К.
У
некоторых
двудольных
(например,
у
свёклы)
в
стеблях
и
корнях
образуются
последовательно
кольцами
к
периферии
дополнительные
слои
К.
(поликамбиальность).
О
К.,
образующем
пробку,
см. Феллоген, Перидерма.
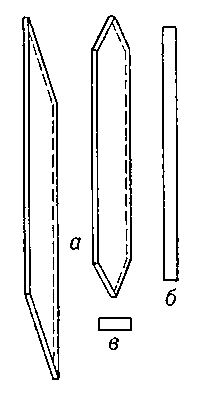
Рис.
1. Схема
прозенхимной
клетки
камбия:
а
—
продольный
тангентальный
разрез;
б
—
продольный
радиальный
разрез;
в
—
поперечный
разрез.
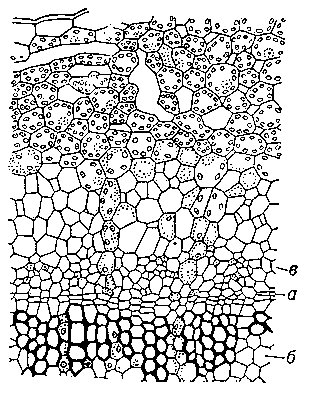
Рис.
2. Часть
поперечного
разреза
стебля
льна:
а
—
камбий;
б
—
древесина;
в
—
луб.
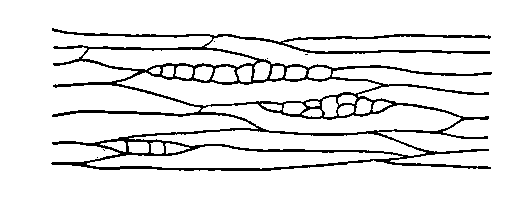
Рис.
3. Клетки
камбия,
разделившиеся
на
клетки
лубо-древесинного
луча.
25)
Хлоре́нхима,
или хлорофилоноснаяпаренхима — ассимиляционная ткань сосудистых
растений,
состоящая
изпаренхимных клеток,
вдоль
тонких
стенок
которых
одним
слоем
располагаются хлоропласты,
не
затеняя
друг
друга.
В
некоторых
руководствах
ассимиляционную
ткань
рассматривают
как
разновидность
основной
паренхимы.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
17.05.201525.76 Mб13Философия. Стёпин.pdf
- #
Источник