
Сосуды растений по которым поступает вода

У этого термина существуют и другие значения, см. Сосуд.
Сосу́ды (трахеи) — проводящие элементы ксилемы, представляющие собой длинные полые трубки, образованные одним рядом клеток (члеников) со сквозными отверстиями (перфорациями) на поперечных стенках, по которым происходит массовое передвижение веществ.
Строение[править | править код]
Сосуды растений (трахеи) состоят из многих клеток, которые называются члениками сосуда. Членики расположены друг над другом, образуя длинную полую трубку. Поперечные перегородки между члениками растворяются, и возникают перфорации (сквозные отверстия). По таким полым трубкам растворы передвигаются значительно легче, чем по трахеидам. Каждый сосуд может состоять из огромного числа члеников, поэтому средняя длина сосудов — несколько сантиметров (иногда до 1 м и больше). Самые совершенные сосуды состоят из широких коротких члеников, диаметр которых превышает длину, а в перфорационных пластинках имеется одно крупное отверстие (простая перфорация). Сосуды менее специализированные состоят из более длинных и узких члеников, поперечные стенки между которыми наклонены. Перфорационные пластинки имеют несколько отверстий, расположенных друг над другом (лестничная перфорация) или в беспорядке (сетчатая перфорация).
Развитие сосуда[править | править код]
Членики сосуда образуются из продольного ряда клеток и вначале представлены расположенными друг над другом живыми паренхимными тонкостенными клетками, полость которых заполнена цитоплазмой с крупным ядром.
Первичная оболочка члеников сосудов состоит из микрофибриллярной фазы и матрикса, заполняющего промежутки между пространственно организованными микрофибриллами целлюлозы. В оболочке молодых члеников сосуда преобладают компоненты матрикса и вода. В связи с этим они могут удлиняться и разрастаться в ширину, протопласт вакуолизируется и занимает постенное положение.
Ещё до завершения роста начинается отложение слоёв вторичной оболочки. Каждый из слоёв отличается направлением ориентации микрофибрилл, характерным для данного типа элементов ксилемы. В тех участках первичной оболочки, где позднее образуются перфорации, вторичная оболочка не откладывается, но за счёт разбухания пектинового вещества межклеточной пластинки эти участки несколько утолщаются.
В самых ранних по времени образования трахеальных элементах вторичная оболочка может иметь форму колец, не связанных друг с другом (кольчатые сосуды). Позднее появляются трахеальные элементы со спиральными утолщениями, затем с лестничными утолщениями (сосуды с утолщениями, которые могут быть охарактеризованы как плотные спирали, витки которых связаны между собой).
Сосуды с относительно небольшими округлыми участками первичной оболочки, не прикрытыми изнутри вторичной оболочкой, называют пористыми.
Вторичная оболочка, а иногда и первичная, как правило, лигнифицируются, то есть пропитываются лигнином. Это придает им дополнительную прочность, но ограничивает возможности дальнейшего роста органа в длину. Одновременно с одревеснением боковых клеток сосуда идет процесс разрушения поперечных стенок между члениками: они ослизняются и постепенно исчезают. Так формируется перфорация. Вокруг перфорации всегда сохраняется остаток продырявленной стенки в виде ободка (перфорационный поясок).
После образования перфорации протопласт отмирает, его остатки в виде бородавчатого слоя выстилают стенки трахеальных элементов (трахеид и члеников сосудов). В результате последовательных структурных изменений формируется сплошная полая трубка сосуда, полость которой заполняется водой.
Механизм действия[править | править код]
Механизм поступления воды в трахеальные элементы и проведения её ко всем частям растения сложен. Основная масса воды поступает в растение через корневые волоски. В силу т. н. корневого давления вода проходит к водопроводящим элементам корня, поднимается к листьям и испаряется с их поверхности наружу через устьица (транспирация).
Сосуды заполнены водой. По мере того, как вода движется по сосудам, в столбе воды создаётся натяжение. Оно передаётся вниз по стеблю на всём пути от листа к корню благодаря сцеплению (когезии) молекул воды. Молекулы стремятся «прилипнуть» друг к другу в силу своей полярности, а затем удерживаются вместе за счёт водородных связей. Кроме того, они стремятся прилипнуть к стенкам сосудов под действием сил адгезии. Натяжение в сосудах ксилемы достигает такой силы, что может тянуть весь столб воды вверх, создавая массовый поток. При этом прочность стенкам обеспечивают целлюлоза и лигнин.
Литература[править | править код]
- Атлас по анатомии растений: учеб. пособие для вузов / Бавтуто Г. А., Ерёмин В. М., Жигар М. П.. — Мн.: Ураджай, 2001. — 146 с. — (Учеб. и учеб. пособия для вузов). — ISBN 985-04-0317-9.
Источник
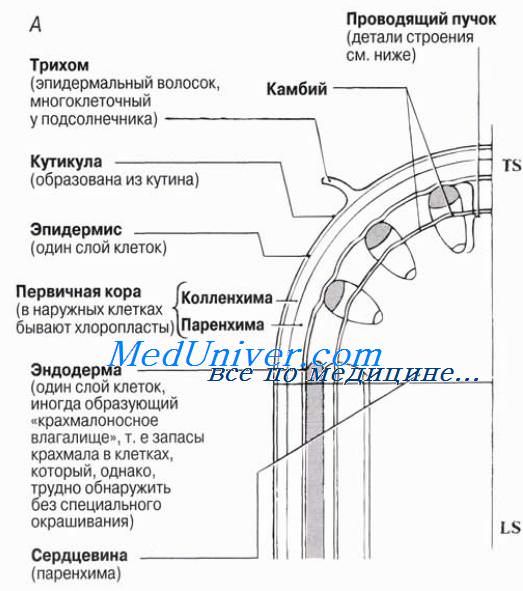
Подъем воды по ксилеме растений.Ксилема цветковых растений содержит два типа проводящих воду структур — трахеиды и сосуды. В статье мы уже говорили о том, как выглядят эти структуры в световом микроскопе, а также на микрофотографиях, полученных с помощью сканирующего электронного микроскопа. Строение вторичной ксилемы (древесины) рассматривается в статье. Ксилема и флоэма образуют проводящую ткань высших, или сосудистых, растений. Эта ткань состоит из так называемых проводящих пучков, строение и распределение которых в стеблях двудольных растений с первичным строением показано на рисунке. То, что вода поднимается именно по ксилеме, нетрудно продемонстрировать, погрузив побег срезанным концом в разбавленный водный раствор красителя, например эозина. Подкрашенная жидкость, распространившись вверх по стеблю, заполнит сеть пронизывающих листья жилок. Если затем сделать тонкие срезы и рассмотреть их в световом микроскопе, то окажется, что краситель находится в ксилеме. Более эффектное доказательство подъема воды по ксилеме дают опыты с «кольцеванием». Такие опыты проводили задолго до того, как стали применяться радиоактивные изотопы, позволяющие очень легко проследить путь веществ в живом организме. В одном из вариантов опыта с одревесневшего стебля снимают узкое кольцо коры вместе с лубом, т. е. флоэмой. Довольно долго после этого находящиеся выше вырезанного кольца побеги продолжают расти нормально: следовательно, такое кольцевание не влияет на подъем воды по стеблю. Однако, если, приподняв лоскут коры, вырезать из-под него сегмент древесины, т. е. ксилемы, то растение быстро завянет. Таким образом, вода движется в побеги из почвы именно по этой проводящей ткани. Любая теория, объясняющая транспорт воды по ксилеме, не может не учитывать следующие наблюдения.
1. Анатомические элементы ксилемы — тонкие мертвые трубки, диаметр которых варьирует от 0,01 мм в «летней» древесине до 0,2 мм в «весенней» древесине. Удовлетворительное объяснение этим фактам дает теория сцепления (когезии), или теория натяжения. Согласно этой теории, подъем воды от корней обусловлен ее испарением клетками листа. Как мы уже говорили в статье, испарение снижает водный потенциал клеток мезофилла, прилежащих к ксилеме, и вода поступает в эти клетки из ксилемного сока, водный потенциал которого выше; при этом она проходит через влажные клеточные стенки у концов жилок, как показано на рисунке. Ксилемные сосуды заполняет сплошной столб воды; по мере того как вода выходит из сосудов, в этом столбе создается натяжение; оно передается вниз по стеблю до самого корня благодаря сцеплению (когезии) молекул воды. Эти молекулы стремятся «прилипнуть» друг к другу, потому что они полярны и притягиваются друг к другу электрическими силами, а затем удерживаются вместе водородными связями. Кроме того, они притягиваются к стенкам ксилемных сосудов, т. е. происходит их адгезия (прилипание) к ним. Сильная когезия молекул воды означает, что ее столб трудно разорвать — у него высокий предел прочности при растяжении. Растягивающее напряжение в клетках ксилемы приводит к генерированию силы, способной сдвигать весь водяной столб вверх по механизму объемного потока. Снизу вода поступает в ксилему из соседних клеток корня. При этом очень важно, что стенки ксилемных элементов жесткие и не спадаются при падении давления внутри, как это бывает, когда сосешь коктейль через мягкую соломинку. Жесткость стенок обеспечивается лигнином. Доказательством того, что жидкость внутри ксилемных сосудов сильно напряжена (растянута), служат суточные колебания диаметра древесных стволов, измеряемые инструментом под названием дендрограф. Минимальный диаметр отмечен днем, когда интенсивность транспирации наивысшая. Натяжение столба воды в ксилемном сосуде немного втягивает внутрь его стенки (из-за адгезии), и сочетание этих микроскопических сжатий дает фиксируемую прибором общую «усадку» ствола. Оценки прочности на разрыв столба ксилемного сока варьировали от 3000 доЗО 000 к Па, причем более низкие значения получены позднее. В листьях зарегистрирован водный потенциал порядка —4000 кПа, и прочность столба ксилемного сока, вероятно, достаточна, чтобы выдержать создающееся натяжение. Не исключено, конечно, что столб воды может иногда разрываться, особенно в сосудах большого диаметра. Критики изложенной теории подчеркивают, что любое нарушение непрерывности столба сока должно немедленно останавливать весь поток, так как сосуд заполнится воздухом и паром (явление кавитации). Кавитацию может вызвать сильное сотрясение, изгибание ствола, а также дефицит воды. Хорошо известно, что на протяжении лета содержание воды в стволе дерева постепенно снижается, древесина заполняется воздухом. Этим пользуются лесозаготовители, потому что такие деревья легче сплавлять. Однако разрыв водного столба в части сосудов слабо влияет на общую скорость объемного потока. Возможно, дело в том, что вода перетекает в параллельно проходящие сосуды или же обходит воздушную пробку, продвигаясь по соседним паренхимным клеткам и по стенкам. Кроме того, согласно расчетам, для поддержания наблюдаемой скорости потока вполне достаточно, чтобы в каждый момент времени функционировала хотя бы небольшая доля ксилемных элементов. У некоторых деревьев и кустарников вода перемещается лишь по более молодой наружной древесине, называемой заболонью. У дуба и ясеня, например, проводящую функцию выполняют в основном сосуды текущего года, а остальная часть заболони играет роль водного резерва. Новые ксилемные сосуды образуются на протяжении всего вегетационного периода, но главным образом в его начале, когда скорость водного потока максимальна. Вторая сила, обеспечивающая движение воды по ксилеме, — корневое давление. Его можно обнаружить и измерить в тот момент, когда срезают крону, а штамб с корнями некоторое время продолжает выделять сок из сосудов ксилемы. Этот процесс подавляется ингибиторами дыхания, например цианидом, и прекращается при недостатке кислорода и понижении температуры. Работа такого механизма, по-видимому, обусловлена активной секрецией солей и других водорастворимых веществ в ксилемный сок. В результате его водный потенциал падает, и вода поступает в ксилему из соседних клеток корня путем осмоса. Этот механизм создает гидростатическое давление порядка 100-200 кПа (в исключительных случаях 800 кПа); одного его для подъема воды по ксилеме обычно недостаточно, однако у многих растений оно, несомненно, способствует поддержанию ксилемного тока. У медленно транспирирующих травянистых форм этого давления вполне хватает, чтобы вызвать у них мутацию. Так называется выделение воды на поверхности растения1 в виде капель жидкости, а не пара. Все условия, тормозящие транспира-цию, например слабая освещенность и высокая влажность, способствуют гуттации. Она обычна у многих видов дождевых тропических лесов и часто наблюдается на кончиках листьев у всходов трав. – Также рекомендуем “Поглощение воды корнями. Апопластный транспорт в корне.” Оглавление темы “Транспорт у растений.”: |
Источник
Хотя представители флоры могут абсорбировать воду всеми частями тела, основным её источником является почва. Но как она попадает от корня к вершине дерева, высотой с десятиэтажный дом? У растений нет мышц, они не могут качать жидкость по системе кровообращения. Тем не менее, вода движется по растениям вверх через клеточные стенки между протопластом клеток, через плазмодесмы (цитоплазматические мостики), через плазматические мембраны, а также через соединённые между собой проводящие элементы, распространённые по всему растению.
Сначала вода входит в корни, потом перемещается по ксилеме – внутренней проводящей ткани растения. Она поднимается, преодолевая силу тяжести Земли, благодаря двум двигателям: нижнему – корневому давлению и верхнему – испарению воды, и покидает растение в парообразном состоянии главным образом через устьица листьев (транспирация).
Большую часть пути наверх молекулы воды проходят по ксилеме. Например, по проводящим путям секвойи раствор может подниматься на высоту до 100 м. Главная роль в передвижении воды вверх принадлежит притягивающей силе, возникающей в результате испарения. При этом молекулы воды сцепляются водородными связями и движутся по стенкам трахеид и сосудов ксилемы (адгезия).
 Части растения, участвующие в подъёме воды
Части растения, участвующие в подъёме воды
Осмос усиливается аквапоринами
Вода в корневую систему поступает в зоне всасывания, через корневые волоски. Механизмы проникновения её в клетки подчиняются общим законом транспорта воды через плазмалемму. Если одиночную клетку поместить в воду, то концентрация ионов внутри ячейки будет больше, чем снаружи неё. И вода станет двигаться в клетку путём осмоса.
Однако скорость осмоса через мембрану ограничена. Долгое время учёные не могли понять, как вода может двигаться быстрее, чем предусматривает скорость осмоса. Теперь мы знаем, что осмос усиливается мембранными водными каналами, которые формируют интегральные мембранные протеины, называемые аквапоринами. Эти каналы есть в клетках животных и растений. Они проходят через мембраны вакуолей и клеточные мембраны и обеспечивают объёмный поток жидкостей.
Мембранные водные каналы ускоряют движение воды по ксилеме, обеспечивают постоянство водного баланса клетки, но они не способны изменить направление потока.

Вода движется по растениям вверх благодаря разности собственного потенциала
Направленное движение воды через плазмалемму обеспечивает разность потенциалов воды в корне и на поверхности устьиц. Потенциал воды – это вид свободной энергии. Именно градиент водного потенциала является решающей силой в определении направления движения жидкости. Жидкость движется от места большей концентрации к тому месту, где она меньше. Потенциал воды измеряется в единицах, называемых мегапаскалями (МПа).
 Корневое питание растения
Корневое питание растения
Как образуется водный потенциал?
Клеточные стенки сдерживают внутреннее давление клетки, когда вода наполняет её. Если клетку поместить в гипертонический раствор (с очень высокой концентрацией сахарозы), вода будет выходить из клетки, а клеточное давление упадёт. Клеточная мембрана отходит от клеточной стенки по мере уменьшения объёма клетки. Когда давление падает до 0, большинство растений вянут.
 Тургор и плазмолиз в клетке растений
Тургор и плазмолиз в клетке растений
Изменение размера тургора можно предсказать путём вычисления потенциала воды в клетке и окружающем растворе. Водный потенциал имеет две составляющие:
- физические силы, такие как гравитация и давление на клеточную стенку;
- концентрация растворённого вещества внутри клетки и снаружи.
Вода всегда движется в направлении более низкого потенциала воды. Например, водопад движется вниз, потому что гравитация является для него основным фактором, а потенциал в нижней части водопада ниже, чем в верхней.
На уровне клетки вклад гравитации в потенциал воды настолько мал, что обычно не входит в расчёты, если не рассматривать очень высокое дерево. Тургорное давление (давление на клеточные стенки) называется потенциалом давления. Как только увеличивается тургор, увеличивается и потенциал давления.
Концентрация растворённых веществ также определяет потенциал воды и называется потенциалом растворённого вещества. В чистой воде он нулевой. Когда в ней растворяют вещества, молекулы воды образуют с ними водородные связи. Становится меньше свободных молекул воды, что уменьшает водный потенциал. Раствор с большей концентрацией веществ имеет меньший потенциал.
Общий водный потенциал растительной клетки – это сумма потенциала её давления и потенциала растворённого вещества. Когда общий потенциал энергии воды внутри и снаружи клетки одинаковый, то вода не движется.

Источник
Появление проводящих тканей в процессе эволюции является одной из причин, которые сделали возможным выход растений на сушу. В нашей статье мы рассмотрим особенности строения и функционирования ее элементов – ситовидных трубок и сосудов.
Особенности проводящей ткани
Когда на планете произошли серьезные изменения климатических условий, растениям пришлось приспосабливаться к ним. До этого все они обитали исключительно в воде. В наземно-воздушной среде стала необходимой добыча воды из почвы и ее транспортировка ко всем органам растения.
Различают два вида проводящей ткани, элементами которой являются сосуды и ситовидные трубки:
- Луб, или флоэма – расположена ближе к поверхности стебля. По ней органические вещества, образованные в листе во время фотосинтеза, передвигаются по направлению к корню.
- Второй тип проводящей ткани называется древесина, или ксилема. Она обеспечивает восходящий ток: от корня к листьям.

Ситовидные трубки растений
Это проводящие клетки луба. Между собой они разделены многочисленными перегородками. Внешне их строение напоминает сито. Отсюда и происходит название. Ситовидные трубки растений живые. Это объясняется слабым давлением нисходящего тока.
Их поперечные стенки пронизаны густой сетью отверстий. А клетки содержат много сквозных отверстий. Все они являются прокариотическими. Это означает, что в них нет оформленного ядра.
Живыми элементы цитоплазмы ситовидных трубок остаются только на определенное время. Продолжительность этого периода варьирует в широких пределах – от 2 до 15 лет. Данный показатель зависит от вида растения и условий его произрастания. Ситовидные трубки транспортируют воду и органические вещества, синтезированные в процессе фотосинтеза от листьев к корню.

Сосуды
В отличие от ситовидных трубок, эти элементы проводящей ткани представляют собой мертвые клетки. Визуально они напоминают трубочки. Сосуды имеют плотные оболочки. С внутренней стороны они образуют утолщения, которые имеют вид колец или спиралей.
Благодаря такому строению сосуды способны выполнять свою функцию. Она заключается в передвижении почвенных растворов минеральных веществ от корня к листьям.
Механизм почвенного питания
Таким образом, в растении одновременно осуществляется передвижение веществ в противоположных направлениях. В ботанике этот процесс называют восходящим и нисходящим током.
Но какие силы заставляют воду из почвы двигаться вверх? Оказывается, что это происходит под влиянием корневого давления и транспирации – испарения воды с поверхности листьев.
Для растений этот процесс является жизненно необходимым. Дело в том, что только в почве находятся минералы, без которых развитие тканей и органов будет невозможным. Так, азот необходим для развития корневой системы. В воздухе этого элемента предостаточно – 75 %. Но растения не способны фиксировать атмосферный азот, поэтому минеральное питание так важно для них.
Поднимаясь, молекулы воды плотно сцепляются между собой и стенками сосудов. При этом возникают силы, способные поднять воду на приличную высоту – до 140 м. Такое давление заставляет почвенные растворы через корневые волоски проникать в кору, и далее к сосудам ксилемы. По ним вода поднимается к стеблю. Далее, под действием транспирации, вода поступает в листья.
В жилках рядом с сосудами находятся и ситовидные трубки. Эти элементы осуществляют нисходящий ток. Под воздействием солнечного света в хлоропластах листа синтезируется полисахарид глюкоза. Это органическое вещество растение расходует на осуществление роста и процессов жизнедеятельности.
Итак, проводящая ткань растения обеспечивает передвижение водных растворов органических и минеральных веществ по растению. Ее структурными элементами являются сосуды и ситовидные трубки.
Источник