
Сосуды состоят из мертвых клеток

Ксилема. Строение ксилемы. Функции ксилемы.Ксилема выполняет в растении две основные функции: по ней движется вода вместе с растворенными минеральными веществами и она служит опорой органам растения. Таким образом, ксилема играет в растении двоякую роль — физиологическую и структурную. В состав ксилемы входят гистологические элементы четырех типов: трахеиды, сосуды, паренхимные клетки и волокна. На рис. 6.9 эти гистологические элементы представлены и поперечном и продольном разрезах. Трахеиды ксилемыТрахеиды — это одиночные лигнифицированные клетки веретеновидной формы. Концы соприкасающихся трахеид перекрываются так же, как и заостренные концы волокон склеренхимы. Это придает трахеидам механическую прочность и обеспечивает органам растения опору. Трахеиды — мертвые клетки; в зрелом состоянии их просвет ничем не заполнен. Среди водопроводя-ших клеток сосудистых растений трахеиды представляют первичную примитивную форму; у древних сосудистых растений это единственные водопроводящие клетки. Из них развились описанные ниже сосуды и волокна ксилемы высших растений. Несмотря на свой примитивный характер, трахеиды, несомненно, функционируют эффективно; об этом свидетельствует тот факт, что у голосеменных растений доставка воды от корней к надземным частям обеспечивается исключительно трахеидами, а ведь большинство голосеменных — древесные породы. Вода движется по пустым просветам трахеид, не встречая на своем пути помех в виде живого содержимого. Из одной трахеиды в другую она переходит либо через поры, через их «замыкающие пленки», либо через нелигнифицированные части клеточных стенок. Характер лигнификации (одревеснения) клеточных стенок трахеид близок к тому, который описан ниже для сосудов. На рисунке представлено строение трахеид. У покрытосеменных число трахеид по сравнению с числом сосудов относительно невелико. Сосуды считаются более эффективным приспособлением для транспорта воды, нежели трахеиды; появление сосудов связано, как полагают, с тем, что у покрытосеменных с их большой листовой поверхностью транспира-ция идет более активно. Сосуды ксилемыСосуды — характерные проводящие элементы ксилемы покрытосеменных. Они представляют собой очень длинные трубки, образовавшиеся в результате слияния ряда клеток, соединившихся «конец в конец». Каждая из клеток, образующих сосуд ксилемы, соответствует трахеиде и называется члеником сосуда. Однако членики сосуда короче и шире трахеид. Первая ксилема, появляющаяся в растении в процессе сто развития, носит название первичной ксилемы; она закладывается у кончика корня и на верхушке побегов. Дифференцированные членики сосудов ксилемы появляются рядами на концах прокамбиальных тяжей. Сосуд возникает, когда соседние членики в данном ряду сливаются в результате разрушения перегородок между ними. Внутри сосуда сохраняются в виде ободков остатки разрушенных торцевых стенок. Слияние члеников сосудов изображено на рисунке.
Протоксилема и метаксилемаПервые по времени образования сосуды — протоксилема — закладываются на верхушке осевых органов, непосредственно под верхушечной меристемой, там, где окружающие их клетки еше продолжают вытягиваться. Зрелые сосуды про-токсилемы способны растягиваться одновременно с вытягиванием окружающих клеток, поскольку их целлюлозные стенки еще не сплошь одревеснели —лигнин откладывается в них лишь кольцами или по спирали (рис. 6.12). Эти отложения лигнина позволяют трубкам сохранять достаточную прочность во время роста стебля или корня. С ростом органа появляются новые сосуды ксилемы, которые претерпевают более интенсивную лигнификацию и завершают свое развитие в зрелых частях органа; так формируется ме-гаксшема. Тем временем самые первые сосуды протоксилемы растягиваются, а затем разрушаются. Зрелые сосуды метаксилемы не способны растягиваться и расти. Это мертвые, жесткие? полностью одревесневшие трубки. Если бы их развитие завершалось до того, как закончилось вытягивание окружающих живых клеток, то они бы очень сильно мешали этому процессу. У сосудов метаксилемы обнаруживаются три главных типа утолщений: лестничные, сетчатые и точечные. Длинные полые трубки ксилемы — идеальная система для проведения воды на большие расстояния с минимальными помехами. Так же как и в трахеидах, вода может переходить из сосуда в сосуд через поры или через неодревеснев-шие части клеточной стенки. Вследствие одревеснения клеточные стенки сосудов обладают высокой прочностью на разрыв, что тоже очень важно, потому что благодаря этому трубки не спадаются, когда вода движется в них под натяжением (разд. 13.4). Вторую свою функцию — механическую — ксилема выполняет также благодаря тому, что она состоит из ряда одревесневших трубок. В первичном теле растения ксилема в корнях занимает центральное положение, помогая корню противостоять тянущему усилию надземных частей, изгибающихся под порывами ветра, В стебле проводящие пучки либо образуют по периферии кольцо, как у двудольных, либо располагаются беспорядочно, как у однодольных; в обоих случаях стебель пронизывается отдельными тяжами ксилемы, обеспечивающими ему определенную опору. Особенно важное значение опорная функция ксилемы приобретает там, где имеет место вторичный рост. Во время этого процесса быстро нарастает количество вторичной ксилемы; к ней переходит от колленхимы и склеренхимы роль главной механической ткани, и именно она служит опорой у крупных древесных и кустарниковых пород. Рост стволов в толщину определяется в известной мере нагрузками, которым подвергается растение, так что иногда наблюдается дополнительный рост, смысл которого состоит в усилении структуры и обеспечении ей максимальной опоры.
Древесинная паренхима ксилемыДревесинная паренхима ксилемы содержится как в первичной, так и во вторичной ксилеме, однако в последней ее количество больше и роль важнее. Клетки древесинной паренхимы, подобно любым другим паренхимным клеткам, имеют тонкие целлюлозные стенки и живое содержимое. Во вторичной ксилеме имеются две системы паренхимы. Обе они возникают из меристемати-ческих клеток, называемых в одном случае лучевыми инициалями, а вдругом — веретеновидны-ми инициалями (гл. 22). Лучевая паренхима более обильна. Она образует радиальные слои ткани, так называемые сердцевинные лучи, которые, пронизывая сердцевину, служат живой связью между сердцевиной и корой. Здесь запасаются различные питательные вещества, скапливаются таннины, кристаллы и т. п., и здесь же осуществляется радиальный транспорт питательных веществ и воды, а также газообмен по межклетникам. Из веретеновидных инициалей обычно развиваются сосуды ксилемы и ситовидные трубки флоэмы вместе с их клетками-спутницами, однако время от времени они дают начало также и паренхимным клеткам. Эти паренхимные клетки образуют во вторичной ксилеме вертикальные ряды. Древесинные волокна ксилемыПолагают, что древесинные волокна, так же как и сосуды ксилемы, ведут свое происхождение от трахеид. Они короче и уже трахеид, а стенки их гораздо толще, но поры их сходны с порами, имеющимися в трахеидах, и на срезах волокна иногда трудно отличить от трахеид, поскольку между теми и другими есть ряд переходных форм. Древесинные волокна очень напоминают уже описанные волокна склеренхимы; их торцевые стенки также перекрываются. В отличие от сосудов ксилемы древесинные волокна не проводят воду; поэтому у них могут быть гораздо более толстые стенки и более узкие просветы, а значит, они отличаются и большей прочностью, т. е. придают ксилеме дополнительную механическую прочность. – Также рекомендуем “Флоэма. Строение флоэмы. Функции флоэмы.” Оглавление темы “Ткани. Питание клетки.”: |
Источник
У этого термина существуют и другие значения, см. Сосуд.
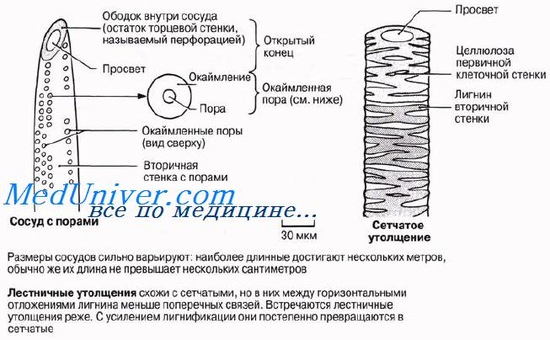
Сосу́ды (трахеи) — проводящие элементы ксилемы, представляющие собой длинные полые трубки, образованные одним рядом клеток (члеников) со сквозными отверстиями (перфорациями) на поперечных стенках, по которым происходит массовое передвижение веществ.
Строение[править | править код]
Сосуды растений (трахеи) состоят из многих клеток, которые называются члениками сосуда. Членики расположены друг над другом, образуя длинную полую трубку. Поперечные перегородки между члениками растворяются, и возникают перфорации (сквозные отверстия). По таким полым трубкам растворы передвигаются значительно легче, чем по трахеидам. Каждый сосуд может состоять из огромного числа члеников, поэтому средняя длина сосудов — несколько сантиметров (иногда до 1 м и больше). Самые совершенные сосуды состоят из широких коротких члеников, диаметр которых превышает длину, а в перфорационных пластинках имеется одно крупное отверстие (простая перфорация). Сосуды менее специализированные состоят из более длинных и узких члеников, поперечные стенки между которыми наклонены. Перфорационные пластинки имеют несколько отверстий, расположенных друг над другом (лестничная перфорация) или в беспорядке (сетчатая перфорация).
Развитие сосуда[править | править код]
Членики сосуда образуются из продольного ряда клеток и вначале представлены расположенными друг над другом живыми паренхимными тонкостенными клетками, полость которых заполнена цитоплазмой с крупным ядром.
Первичная оболочка члеников сосудов состоит из микрофибриллярной фазы и матрикса, заполняющего промежутки между пространственно организованными микрофибриллами целлюлозы. В оболочке молодых члеников сосуда преобладают компоненты матрикса и вода. В связи с этим они могут удлиняться и разрастаться в ширину, протопласт вакуолизируется и занимает постенное положение.
Ещё до завершения роста начинается отложение слоёв вторичной оболочки. Каждый из слоёв отличается направлением ориентации микрофибрилл, характерным для данного типа элементов ксилемы. В тех участках первичной оболочки, где позднее образуются перфорации, вторичная оболочка не откладывается, но за счёт разбухания пектинового вещества межклеточной пластинки эти участки несколько утолщаются.
В самых ранних по времени образования трахеальных элементах вторичная оболочка может иметь форму колец, не связанных друг с другом (кольчатые сосуды). Позднее появляются трахеальные элементы со спиральными утолщениями, затем с лестничными утолщениями (сосуды с утолщениями, которые могут быть охарактеризованы как плотные спирали, витки которых связаны между собой).
Сосуды с относительно небольшими округлыми участками первичной оболочки, не прикрытыми изнутри вторичной оболочкой, называют пористыми.
Вторичная оболочка, а иногда и первичная, как правило, лигнифицируются, то есть пропитываются лигнином. Это придает им дополнительную прочность, но ограничивает возможности дальнейшего роста органа в длину. Одновременно с одревеснением боковых клеток сосуда идет процесс разрушения поперечных стенок между члениками: они ослизняются и постепенно исчезают. Так формируется перфорация. Вокруг перфорации всегда сохраняется остаток продырявленной стенки в виде ободка (перфорационный поясок).
После образования перфорации протопласт отмирает, его остатки в виде бородавчатого слоя выстилают стенки трахеальных элементов (трахеид и члеников сосудов). В результате последовательных структурных изменений формируется сплошная полая трубка сосуда, полость которой заполняется водой.
Механизм действия[править | править код]
Механизм поступления воды в трахеальные элементы и проведения её ко всем частям растения сложен. Основная масса воды поступает в растение через корневые волоски. В силу т. н. корневого давления вода проходит к водопроводящим элементам корня, поднимается к листьям и испаряется с их поверхности наружу через устьица (транспирация).
Сосуды заполнены водой. По мере того, как вода движется по сосудам, в столбе воды создаётся натяжение. Оно передаётся вниз по стеблю на всём пути от листа к корню благодаря сцеплению (когезии) молекул воды. Молекулы стремятся «прилипнуть» друг к другу в силу своей полярности, а затем удерживаются вместе за счёт водородных связей. Кроме того, они стремятся прилипнуть к стенкам сосудов под действием сил адгезии. Натяжение в сосудах ксилемы достигает такой силы, что может тянуть весь столб воды вверх, создавая массовый поток. При этом прочность стенкам обеспечивают целлюлоза и лигнин.
Литература[править | править код]
- Атлас по анатомии растений: учеб. пособие для вузов / Бавтуто Г. А., Ерёмин В. М., Жигар М. П.. — Мн.: Ураджай, 2001. — 146 с. — (Учеб. и учеб. пособия для вузов). — ISBN 985-04-0317-9.
Источник
Трахеальные элементы представляют
собой наиболее высокоспециализированные клетки
ксилемы. Как правило, они вытянуты в длину и в
зрелом состоянии мертвы. Для них характерны
лигнифицированные оболочки со вторичными
утолщениями и порами.
Процесс отложения вторичной оболочки
и пропитывание ее лигнином осуществляется еще в
живой клетке. При этом на первых этапах развития
растения все клетки растущих частей удлиняются
(вытягиваются). Однако такое удлинение было бы
невозможным при сплошной жесткой оболочке. В
этом отношении у высших растений выработалось
оптимальное приспособление: вторичная оболочка
не одевает клетку сплошь, а расположена кольцами
или спиралью. Подобные кольчатые и спиральные
утолщения позволяют молодым трахеальным
элементам вытягиваться в длину и в то же время
препятствуют из сдавливанию. Кроме того, этот тип
вторичного утолщения является чрезвычайно
экономичным.
При всех достоинствах кольчатых и
спиральных элементов как путей для проведения
воды, их механическая прочность оставляет желать
лучшего. Поэтому, как только у молодого растения
заканчиваются ростовые процессы, связанные с
удлинением клеток, в ксилеме начинают
формироваться трахеальные элементы со сплошной
вторичной одревесневшей оболочкой. Когда
формирование сплошной оболочки заканчивается,
клетки очень быстро отмирают.
Однако и в этом случае оболочка
водопроводящего элемента не может быть
совершенно сплошной. Обычно в ней имеются
многочисленные углубления в виде пор. Поэтому
зрелые водопровдящие элементы называют
точечно-поровыми. Итак, в процессе онтогенеза
(индивидуального развития растения) наблюдается
ряд взаимопревращения трахеальных элементов:
кольчатые, спиральные, сетчатые, лестничные,
точечно-поровые.
Различают два типа проводящих
элементов: 1) трахеиды и 2) членики сосудов.
Отличаются они главным образом тем, что членики
сосудов имеют сквозные отверстия ≈ перфорации, в
то время как трахеиды являются
неперфорированными элементами. Поэтому по
сосудам растворы продвигаются значительно
легче, чем по трахеидам.
Трахеиды ≈ это основной
водопроводящий элемент высших растений с
момента их появления на суше и по сей день.
Трахеиды имеют замкнутую со всех сторон
первичную оболочку. Поэтому вода по трахеидам
должна проходить через окаймленные поры,
просачиваясь через первичные оболочки и
склеивающий их межклеточный слой. Понятно, что
подобная структура не является оптимальной для
быстрой подачи воды.
Поэтому в процессе эволюции в ряде
групп высших растений возник новый более
совершенный проводящий элемент ≈ членик сосуда.
Для члеников сосудов характерно наличие в концах
клеток перфораций. Соединяясь между собой
перфорациями, сотни и тысячи члеников образуют
сосуды ≈ длинные сквозные трубки, по которым
вода перемещается практически беспрепятственно.
Водопроводящие элементы, обладающие
достаточно толстой оболочкой, инкрустированной
лигнином, играют в теле растения также и
механическую роль.
Поэтому специализация трахеальных
элементов в процессе эволюции шла в направлении
разделения механической и проводящей функций.
Примитивная древесина большинства
хвощей, папоротников, голосеменных имеет гомогенную
(однородную) ксилему и состоит исключительно из
трахеид и небольшого количества древесинной
паренхимы. В такой древесине узкопросветные
толстостенные трахеиды выполняют
преимущественно механическую функцию, а
широкопросветные и тонкостенные ≈
водопроводящую.
Более совершенная гетерогенная
древесина покрытосеменных состоит из сосудов,
трахеид, волокон – либриформа и запасающей
паренхимы.
Членики сосудов у цветковых растений
морфологически довольно разнообразны и образуют
четкий эволюционный ряд. Этот эволюционный ряд,
построенный на работах выдающегося
американского ботаника И. Бэйли, помещается во
всех учебниках, как одна из ярких и бесспорных
демонстраций эволюционных преобразований.
Эволюционный ряд члеников сосудов
начинается с длинных трахеид, имеющих лестничную
поровость и косые заостренные концы. Постепенно
клетки укорачиваются, становятся более широкими,
а их концевые стенки все менее наклонными и в
конце концов оказываются поперечными.
В наиболее примитивных члениках
лестничная перфорационная пластинка имеет
множество перегородок. В процессе эволюции они
сокращаются и исчезают совсем, так что остается
одна сквозная крупная перфорация.
Характерно, что с появлением сосудов
цветковых растений не исчезли трахеиды,
поскольку совершенный водопроводящий аппарат не
всегда является преимуществом. Так, в условиях
высокой влажности и затенения способность
быстро проводить воду не имеет особого значения.
У таких растений процент трахеид и примитивных
члеников сосудов с лестничной перфорацией
заметно выше, чем у растений, произрастающих в
засушливых условиях.
Получается, что соотношение
проводящих элементов четко обусловлено
экологическими условиями и определяет водный
баланс растения.
Мы уже говорили, что сочетание в одном
клеточном элементе ≈ трахеиде ≈ и способности
проводить воду и механической прочности было
важным эволюционным приспособлением. Появление
сосудов, с их все увеличивающейся полостью и
относительно тонкими стенками, несколько
ослабило бы механическую роль ксилемы, если
сосуды остались бы единственным элементом этой
ткани. В связи с этим наметился второй
эволюционный ряд специализации в сторону
повышения механической прочности, и трахеиды,
через промежуточную стадию волокнистой трахеиды
превратились в волокна либриформа.
При этом происходило утолщение
оболочек клеток, сужение полостей, все большая
редукция окаймления пор. Поры у волокон
либриформа стали узкие, щелевидные. Вместе с тем,
сократилось и количество пор.
Паренхимные клетки, входящие в состав
древесины, запасают крахмал, жиры и многие другие
эргастические вещества.
В ходе онтогенеза у растения из
первичной латеральной меристемы прокамбия
появляется первичная ксилема. У некоторых
растений со временем начинает работать
вторичная боковая меристема ≈ камбий, дающая
начало вторичной ксилеме.
Обычно первичная ксилема отчетливо
подразделяется на два структурных типа: 1) протоксилему
и 2) метаксилему
Источник