
Сосуды стенки клеток толстые одревесневшие

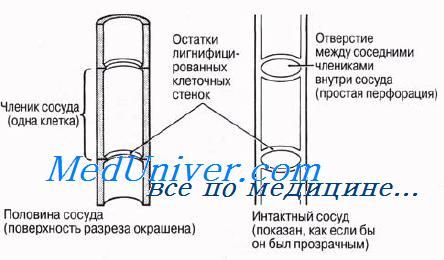
Ксилема. Строение ксилемы. Функции ксилемы.Ксилема выполняет в растении две основные функции: по ней движется вода вместе с растворенными минеральными веществами и она служит опорой органам растения. Таким образом, ксилема играет в растении двоякую роль — физиологическую и структурную. В состав ксилемы входят гистологические элементы четырех типов: трахеиды, сосуды, паренхимные клетки и волокна. На рис. 6.9 эти гистологические элементы представлены и поперечном и продольном разрезах. Трахеиды ксилемыТрахеиды — это одиночные лигнифицированные клетки веретеновидной формы. Концы соприкасающихся трахеид перекрываются так же, как и заостренные концы волокон склеренхимы. Это придает трахеидам механическую прочность и обеспечивает органам растения опору. Трахеиды — мертвые клетки; в зрелом состоянии их просвет ничем не заполнен. Среди водопроводя-ших клеток сосудистых растений трахеиды представляют первичную примитивную форму; у древних сосудистых растений это единственные водопроводящие клетки. Из них развились описанные ниже сосуды и волокна ксилемы высших растений. Несмотря на свой примитивный характер, трахеиды, несомненно, функционируют эффективно; об этом свидетельствует тот факт, что у голосеменных растений доставка воды от корней к надземным частям обеспечивается исключительно трахеидами, а ведь большинство голосеменных — древесные породы. Вода движется по пустым просветам трахеид, не встречая на своем пути помех в виде живого содержимого. Из одной трахеиды в другую она переходит либо через поры, через их «замыкающие пленки», либо через нелигнифицированные части клеточных стенок. Характер лигнификации (одревеснения) клеточных стенок трахеид близок к тому, который описан ниже для сосудов. На рисунке представлено строение трахеид. У покрытосеменных число трахеид по сравнению с числом сосудов относительно невелико. Сосуды считаются более эффективным приспособлением для транспорта воды, нежели трахеиды; появление сосудов связано, как полагают, с тем, что у покрытосеменных с их большой листовой поверхностью транспира-ция идет более активно. Сосуды ксилемыСосуды — характерные проводящие элементы ксилемы покрытосеменных. Они представляют собой очень длинные трубки, образовавшиеся в результате слияния ряда клеток, соединившихся «конец в конец». Каждая из клеток, образующих сосуд ксилемы, соответствует трахеиде и называется члеником сосуда. Однако членики сосуда короче и шире трахеид. Первая ксилема, появляющаяся в растении в процессе сто развития, носит название первичной ксилемы; она закладывается у кончика корня и на верхушке побегов. Дифференцированные членики сосудов ксилемы появляются рядами на концах прокамбиальных тяжей. Сосуд возникает, когда соседние членики в данном ряду сливаются в результате разрушения перегородок между ними. Внутри сосуда сохраняются в виде ободков остатки разрушенных торцевых стенок. Слияние члеников сосудов изображено на рисунке.
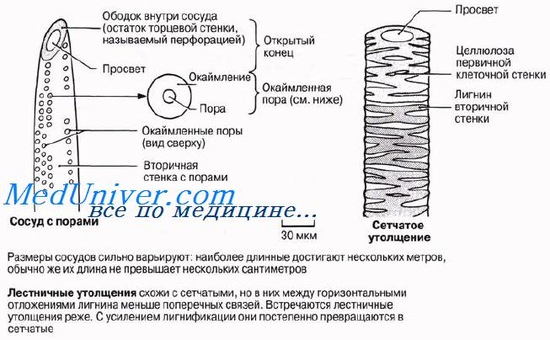
Протоксилема и метаксилемаПервые по времени образования сосуды — протоксилема — закладываются на верхушке осевых органов, непосредственно под верхушечной меристемой, там, где окружающие их клетки еше продолжают вытягиваться. Зрелые сосуды про-токсилемы способны растягиваться одновременно с вытягиванием окружающих клеток, поскольку их целлюлозные стенки еще не сплошь одревеснели —лигнин откладывается в них лишь кольцами или по спирали (рис. 6.12). Эти отложения лигнина позволяют трубкам сохранять достаточную прочность во время роста стебля или корня. С ростом органа появляются новые сосуды ксилемы, которые претерпевают более интенсивную лигнификацию и завершают свое развитие в зрелых частях органа; так формируется ме-гаксшема. Тем временем самые первые сосуды протоксилемы растягиваются, а затем разрушаются. Зрелые сосуды метаксилемы не способны растягиваться и расти. Это мертвые, жесткие? полностью одревесневшие трубки. Если бы их развитие завершалось до того, как закончилось вытягивание окружающих живых клеток, то они бы очень сильно мешали этому процессу. У сосудов метаксилемы обнаруживаются три главных типа утолщений: лестничные, сетчатые и точечные. Длинные полые трубки ксилемы — идеальная система для проведения воды на большие расстояния с минимальными помехами. Так же как и в трахеидах, вода может переходить из сосуда в сосуд через поры или через неодревеснев-шие части клеточной стенки. Вследствие одревеснения клеточные стенки сосудов обладают высокой прочностью на разрыв, что тоже очень важно, потому что благодаря этому трубки не спадаются, когда вода движется в них под натяжением (разд. 13.4). Вторую свою функцию — механическую — ксилема выполняет также благодаря тому, что она состоит из ряда одревесневших трубок. В первичном теле растения ксилема в корнях занимает центральное положение, помогая корню противостоять тянущему усилию надземных частей, изгибающихся под порывами ветра, В стебле проводящие пучки либо образуют по периферии кольцо, как у двудольных, либо располагаются беспорядочно, как у однодольных; в обоих случаях стебель пронизывается отдельными тяжами ксилемы, обеспечивающими ему определенную опору. Особенно важное значение опорная функция ксилемы приобретает там, где имеет место вторичный рост. Во время этого процесса быстро нарастает количество вторичной ксилемы; к ней переходит от колленхимы и склеренхимы роль главной механической ткани, и именно она служит опорой у крупных древесных и кустарниковых пород. Рост стволов в толщину определяется в известной мере нагрузками, которым подвергается растение, так что иногда наблюдается дополнительный рост, смысл которого состоит в усилении структуры и обеспечении ей максимальной опоры.
Древесинная паренхима ксилемыДревесинная паренхима ксилемы содержится как в первичной, так и во вторичной ксилеме, однако в последней ее количество больше и роль важнее. Клетки древесинной паренхимы, подобно любым другим паренхимным клеткам, имеют тонкие целлюлозные стенки и живое содержимое. Во вторичной ксилеме имеются две системы паренхимы. Обе они возникают из меристемати-ческих клеток, называемых в одном случае лучевыми инициалями, а вдругом — веретеновидны-ми инициалями (гл. 22). Лучевая паренхима более обильна. Она образует радиальные слои ткани, так называемые сердцевинные лучи, которые, пронизывая сердцевину, служат живой связью между сердцевиной и корой. Здесь запасаются различные питательные вещества, скапливаются таннины, кристаллы и т. п., и здесь же осуществляется радиальный транспорт питательных веществ и воды, а также газообмен по межклетникам. Из веретеновидных инициалей обычно развиваются сосуды ксилемы и ситовидные трубки флоэмы вместе с их клетками-спутницами, однако время от времени они дают начало также и паренхимным клеткам. Эти паренхимные клетки образуют во вторичной ксилеме вертикальные ряды. Древесинные волокна ксилемыПолагают, что древесинные волокна, так же как и сосуды ксилемы, ведут свое происхождение от трахеид. Они короче и уже трахеид, а стенки их гораздо толще, но поры их сходны с порами, имеющимися в трахеидах, и на срезах волокна иногда трудно отличить от трахеид, поскольку между теми и другими есть ряд переходных форм. Древесинные волокна очень напоминают уже описанные волокна склеренхимы; их торцевые стенки также перекрываются. В отличие от сосудов ксилемы древесинные волокна не проводят воду; поэтому у них могут быть гораздо более толстые стенки и более узкие просветы, а значит, они отличаются и большей прочностью, т. е. придают ксилеме дополнительную механическую прочность. – Также рекомендуем “Флоэма. Строение флоэмы. Функции флоэмы.” Оглавление темы “Ткани. Питание клетки.”: |
Источник
Вены. Строение вен. Стенки и структура вен.Вены в целом сходны по строению с артериями, однако особенности гемодинамики (низкое давление и медленное движение крови в венах) придают структуре их стенки ряд особенностей. По сравнению с артериями одноименные вены имеют больший диаметр (в венозном звене сосудистого русла находится около 70% всей крови), тонкую, легко спадающуюся стенку, слабо развитый эластический компонент, более слабо развитые гладкомышечные элементы в средней оболочке, хорошо выраженную наружную оболочку. Вены, расположенные ниже уровня сердца, имеют полулунные клапаны. Границы между оболочками в венах менее отчетливы по сравнению с артериями. Внутренняя оболочка вен состоит из эндотелия и подэндотелиального слоя. Внутренняя эластическая мембрана слабо выражена. Средняя оболочка вен представлена гладкими мышечными клетками, которые не образуют сплошного слоя, как в артериях, а располагаются в виде обособленных пучков, отделенных прослойками волокнистой соединительной ткани. Эластических волокон мало. Наружная адвентициальная оболочка представляет собой наиболее толстый слой стенки вены. Она содержит коллагеновые и эластические волокна, сосуды, питающие вену, и нервные элементы. Толстая адвентиция вен, как правило, непосредственно переходит в окружающую рыхлую соединительную ткань и фиксирует вену в соседних тканях.
В зависимости от степени развития мышечных элементов вены подразделяются на безмышечные и мышечные. Безмышечные вены располагаются в участках органов с плотными стенками (твердая мозговая оболочка, кости, трабекулы селезенки), в сетчатке глаза, плаценте. В костях и трабекулах селезенки, например, стенки вен сращены своей наружной оболочкой с интерстициальной тканью органов и, таким образом, не спадаются. Строение стенки вен безмышечного типа достаточно простое — эндотелий, окруженный слоем рыхлой соединительной ткани. Гладкомышечных клеток в стенке нет. В венах мышечного типа гладкомышечные клетки имеются во всех трех оболочках. Во внутренней и наружной оболочках пучки гладких миоцитов имеют продольное направление, в средней — циркулярное. Мышечные вены подразделяются на несколько видов. Вены со слабым развитием мышечных элементов — это мелкие вены верхней части туловища, по которым кровь движется, главным образом, вследствие собственной силы тяжести; вены со средним развитием мышечных элементов (мелкие вены, плечевая, верхняя полая вены). В составе внутренней и наружной оболочек этих вен присутствуют единичные продольно ориентированные пучки гладкомышечных клеток, а в средней оболочке — циркулярные пучки гладких миоцитов, разделенные рыхлой соединительной тканью. Эластических мембран в структуре стенки нет, а внутренняя оболочка по ходу вены образует немногочисленные полулунные складки — клапаны, свободные края которых направлены к сердцу. В основании клапанов находятся эластические волокна и гладкомышечные клетки. Предназначение клапанов — препятствовать обратному току крови под влиянием ее собственной силы тяжести. Клапаны открываются по ходу кровотока. Наполняясь кровью, они перекрывают просвет вены и препятствуют обратному движению крови. – Также рекомендуем “Лимфатические сосуды. Строение лимфатических сосудов. Стенки лимфатических сосудов.” Оглавление темы “Сердечно-сосудистая система. Дыхательная система.”: |
Источник
13. Основные ткани: особенности строения клеток, классификация.
Основные ткани составляют основную массу тела растения. Они состоят из живых, относительно мало специализированных клеток, чаще паренхимной формы, поэтому их часто называют паренхимными тканями, или паренхимой. В зависимости от выполняемой функции, различают несколько типов основных тканей.
Ассимиляционная ткань (хлорофиллоносная паренхима, хлоренхима) выполняет функцию фотосинтеза. Она располагается в основном в листьях и стеблях травянистых растений сразу за эпидермой. Клетки живые, тонкостенные, чаще паренхимной формы. 70-80% объема протопласта составляют хлоропласты. Характерно наличие межклетников, которые облегчают газообмен.
Запасающая паренхима служит местом отложения питательных веществ (крахмала, белков, жирных масел). Запасные питательные вещества могут откладываться в живых клетках любой ткани, но особенно ярко эта функция проявляется у специализированных запасающих тканей, хорошо развитых в семенах, корнях, подземных побегах. Состоят запасающие ткани из живых тонкостенных клеток, чаще паренхимной формы.
Разновидностью запасающей ткани является водоносная паренхима, выполняющая функцию запасания воды. Она состоит из крупных живых тонкостенных клеток, как правило, паренхимной формы. Вода запасается в вакуолях за счет большого содержания слизей, обладающих высокой водоудерживающей способностью. Водоносная паренхима имеется в стеблях и листьях суккулентов (кактусы, агавы, алоэ), у многих растений солончаков (солерос, анабазис, саксаул), в листьях многих злаков. Много воды содержится в запасающих тканях луковиц и клубней.
Воздухоносная паренхима (аэренхима) выполняет функцию вентиляции, снабжая ткани и органы кислородом. Она хорошо развита в погруженных органах водных и болотных растений (кувшинка, кубышка, аир, вахта). Аэренхима состоит из живых клеток различной формы и крупных межклетников
14. Ассимиляционная ткань: функция, разновидности, локализация.
Мезофилл занимает все пространство между верхней и нижней эпидермой, исключая проводящие и механические ткани. Клетки мезофилла довольно однородны, по форме чаще всего округлые или слегка вытянутые. Клеточные стенки остаются тонкими и неодревесневшими. Протопласт состоит из постенного слоя цитоплазмы с ядром и многочисленными хлоропластами. В центре клетки находится крупная вакуоль. Иногда стенки клеток образуют складки, которые увеличивают поверхность постенного слоя цитоплазмы и позволяют разместить большее число хлоропластов.
У большинства растений мезофилл дифференцирован на палисадную (столбчатую) и губчатую ткани.
Клетки палисадного мезофилла, расположенного, как правило, под верхней эпидермой, вытянуты перпендикулярно поверхности листа и образуют один или несколько слоев. Клетки губчатого мезофилла соединены более рыхло, межклетные пространства здесь могут быть очень большими по сравнению с объемом самих клеток. Увеличение межклетных пространств часто достигается тем, что клетки губчатого мезофилла образуют выросты.
Палисадная ткань содержит примерно три четверти всех хлоропластов листа и выполняет главную работу по ассимиляции углекислого газа. Поэтому палисадная ткань располагается в наилучших условиях освещения, непосредственно под верхней эпидермой. Благодаря тому, что клетки вытянуты перпендикулярно поверхности листа, лучи света легче проникают вглубь мезофилла.
Через губчатый мезофилл происходит газообмен. Углекислый газ из атмосферы через устьица, расположенные главным образом в нижней эпидерме, проникает в большие межклетники губчатого мезофилла и свободно расходится внутри листа. Кислород, выделяемый при фотосинтезе, передвигается в обратном направлении и через устьица выходит в атмосферу. Расположение устьиц преимущественно на нижней стороне листа объясняется не только положением губчатого мезофилла. Потеря воды листом в процессе транспирации идет медленнее через устьица, расположенные в нижней эпидерме. Кроме того, главным источником углекислого газа в атмосфере является «почвенное дыхание», т. е. выделение СО2 в результате дыхания многочисленных живых существ, населяющих почву.
15. Механические ткани: классификация, функции, особенности строения клеток.
Механические (скелетные, опорные, арматурные) ткани выполняют в растении роль скелета, который скрепляет ткани и части органов между собой. Они придают растениям прочность, способность противостоять действию тяжести собственных органов, порывам ветра, дождю, снегу, вытаптыванию животными. Клетки механических тканей разнообразны по форме, но имеют общий признак – сильно утолщенные клеточные стенки, которые даже после отмирания протопласта продолжают выполнять опорную функцию. Различают два типа механических тканей: 1) колленхиму и 2) склеренхиму.
Колленхима – механическая ткань молодых растущих органов, возникает очень рано, когда еще продолжается рост органа в длину. Колленхима состоит из живых, вытянутых по оси органа клеток с тупыми или скошенными концами. В клетках часто содержатся хлоропласты. Клеточные стенки утолщены неравномерно и никогда не одревесневают. Колленхима располагается сразу за покровной тканью в молодых стеблях, цветоносах, черешках листьев, образуя сплошной цилиндр или тяжи в ребрах. В корнях колленхима обычно не встречается. Наиболее характерна колленхима для двудольных, у однодольных растений она встречается редко.
В зависимости от характера утолщения стенок клеток различают три типа колленхимы: 1) уголковую, 2) пластинчатую и 3) рыхлую.
Склеренхима встречается наиболее часто, во всех органах: корнях, стеблях, листьях, плодах, цветках, семенах. Клетки склеренхимы имеют равномерно утолщенные и, как правило, одревесневшие стенки. Полость клетки мала, поры простые, щелевидные, немногочисленные. Протопласт, как правило, рано отмирает, и опорную функцию выполняют мертвые клетки. Различают два типа склеренхимы: 1) волокна и 2) склереиды, различающиеся формой клеток.
Волокна – прозенхимные клетки, сильно вытянутые в длину и заостренные на концах. Они обеспечивают прочность органов растений на растяжение, сжатие и изгибы. Прочность волокон повышается благодаря тому, что фибриллы целлюлозы проходят в них винтообразно, меняя направление во внешних и внутренних витках.
Волокна, расположенные в коровой части осевого органа (во флоэме), называются лубяными. Лубяные волокна с неодревесневающей клеточной стенкой являются ценным сырьем для текстильной промышленности (лен, рами, кенаф). Волокна, находящиеся в древесине (ксилеме), называются древесинными, или волокнами либриформа. Их стенки всегда одревесневшие, длина их не превышает 2 мм.
Склереиды – клетки, имеющие различную форму, чаще паренхимную. Они встречаются как поодиночке, в виде идиобластов, так и группами. Это мертвые клетки с очень толстыми одревесневшими стенками, пронизанными поровыми каналами, которые часто ветвятся.
16. Проводящие ткани: классификация, функции, особенности строения.
Проводящие элементы ксилемы носят название трахеальных элементов. Различают два типа трахеальных элементов – трахеиды и членики сосудов.
Трахеида представляет собой сильно вытянутую в длину клетку с ненарушенными первичными стенками. Передвижение растворов происходит путем фильтрации через окаймленные поры. Сосуд состоит из многих клеток, называемых члениками сосуда. Членики расположены друг над другом, образуя трубочку. Между соседними члениками одного и того же сосуда имеются сквозные отверстия – перфорации. По сосудам растворы передвигаются значительно легче, чем по трахеидам.
Трахеальные элементы в зрелом, функционирующем состоянии – мертвые клетки, не имеющие протопластов. Сохранение протопластов затрудняло бы передвижение растворов.
Сосуды и трахеиды передают растворы не только в вертикальном, но и в горизонтальном направлении в соседние трахеальные элементы и в живые клетки. Боковые стенки трахеид и сосудов сохраняются тонкими на большей или меньшей площади. В то же время они имеют вторичные утолщения, придающие стенкам прочность. В зависимости от характера утолщений боковых стенок трахеальные элементы называются кольчатыми, спиральными, сетчатыми, лестничными и точечно-поровыми
Ситовидные клетки являются более примитивными, они присущи споровым и голосеменным растениям. Ситовидная клетка – это одна клетка, сильно вытянутая в длину, с заостренными концами. Ее ситовидные поля рассеяны по боковым стенкам. Кроме того, ситовидные клетки имеют и другие примитивные признаки: они лишены специализированных сопровождающих клеток и в зрелом состоянии содержат ядра.
У покрытосеменных растений транспорт ассимилятов осуществляют ситовидные трубки. Они состоят из многих отдельных клеток – члеников, расположенных один над другим. Ситовидные поля двух соседних члеников образуют ситовидную пластинку. Ситовидные пластинки имеют более совершенное строение, чем ситовидные поля (перфорации крупнее и их больше).
В члениках ситовидных трубок в зрелом состоянии отсутствуют ядра, однако они остаются живыми и деятельно проводят вещества. Важная роль в проведении ассимилятов по ситовидным трубкам принадлежит сопровождающим клеткам (клеткам-спутницам). Клетки–спутницы имеют ядра и цитоплазму с многочисленными митохондриями; в них происходит интенсивный обмен веществ. Между ситовидными трубками и прилегающими к ним сопровождающими клетками имеются многочисленные цитоплазматические связи. Считается, что клетки-спутницы вместе с члениками ситовидных трубок составляют единую физиологическую систему, осуществляющую ток ассимилятов.
17. Ксилема, флоэма: функции, состав, особенности строения.
В связи с жизнью в двух средах – почвенной и воздушной, возникли две проводящие ткани, по которым вещества передвигаются в двух направлениях. По ксилеме от корней к листьям поднимаются вещества почвенного питания – вода и растворенные в ней минеральные соли (восходящий, или транспирационный ток). По флоэме от листьев к корням передвигаются вещества, образовавшиеся в процессе фотосинтеза, главным образом сахароза (нисходящий ток). Так как эти вещества представляют собой продукты ассимиляции углекислого газа, транспорт веще ств по флоэме называют током ассимилятов.
Ксилема и флоэма представляют собой сложные ткани, в их состав входят разнородные элементы – проводящие, механические, запасающие, выделительные. Самыми важными являются проводящие элементы, именно они выполняют функцию проведения веществ.
Ксилема (древесина) состоит из проводящих элементов – трахеид и сосудов (трахей), механических элементов – древесинных волокон (волокон либриформа) и элементов основной ткани – древесинной паренхимы.
Флоэма (луб) состоит из проводящих – ситовидных – элементов, сопровождающих клеток (клеток-спутниц), механических элементов – флоэмных (лубяных) волокон и элементов основной ткани – флоэмной (лубяной) паренхимы.
В отличие от трахеальных элементов проводящие элементы флоэмы и в зрелом состоянии остаются живыми, а их клеточные стенки – первичными, неодревесневшими. На стенках ситовидных элементов имеются группы мелких сквозных отверстий – ситовидные поля, через которые сообщаются протопласты соседних клеток и происходит транспорт веществ. Различают два типа ситовидных элементов – ситовидные клетки и членики ситовидных трубок.
В теле растения ксилема и флоэма расположены рядом, образуя или слои, или обособленные тяжи, которые называют проводящими пучками. Различают несколько типов проводящих пучков.
Закрытые пучки состоят только из первичных проводящих тканей, они не имеют камбия и далее не утолщаются. Закрытые пучки характерны для споровых и однодольных растений. Открытые пучки имеют камбий и способны к вторичному утолщению. Они характерны для голосеменных и двудольных растений.
В зависимости от взаимного расположения флоэмы и ксилемы в пучке различают следующие типы. Наиболее обычны коллатеральные пучки, в которых флоэма лежит по одну сторону от ксилемы. Коллатеральные пучки могут быть открытыми (стебли двудольных и голосеменных растений) и закрытыми (стебли однодольных растений). Если с внутренней стороны от ксилемы располагается дополнительно тяж флоэмы, такой пучок называется биколлатеральным. Биколлатеральные пучки могут быть только открытыми, они характерны для некоторых семейств двудольных растений (тыквенные, пасленовые и др.).
Встречаются также концентрические пучки, в которых одна проводящая ткань окружает другую. Они могут быть только закрытыми. Если в центре пучка находится флоэма, а ксилема ее окружает, пучок называется центрофлоэмным, или амфивазальным. Такие пучки часто встречаются в стеблях и корневищах однодольных растений. Если в центре пучка располагается ксилема, и ее окружает флоэма, пучок называется центроксилемным, или амфикрибральным.
18. Корень: определение, функции, корневые системы.
Корень – основной вегетативный орган растения, выполняющий в типичном случае функцию почвенного питания. Корень – осевой орган, обладающий радиальной симметрией и неопределенно долго нарастающий в длину благодаря деятельности апикальной меристемы. От побега он морфологически отличается тем, что на нем никогда не образуются листья, а апикальная меристема всегда прикрыта корневым чехликом.
Кроме главной функции поглощения веществ из почвы, корни выполняют и другие функции:
1) корни укрепляют («заякоривают») растения в почве, делают возможным вертикальный рост и вынесение побегов наверх;
2) в корнях синтезируются различные вещества, которые затем передвигаются в другие органы растения;
3) в корнях могут откладываться запасные вещества;
4) корни взаимодействуют с корнями других растений, микроорганизмами, грибами, обитающими в почве.
Совокупность корней одной особи образует единую в морфологическом и физиологическом отношении корневую систему.
В состав корневых систем входят корни различной морфологической природы – главный корень, боковые и придаточные корни.
Главный корень развивается из зародышевого корешка. Боковые корни образуются на корне (главном, боковом, придаточном), который по отношению к ним обозначается как материнский. Они возникают на некотором расстоянии от апекса, в направлении от основания корня к его верхушке. Боковые корни закладываются эндогенно, т.е. во внутренних тканях материнского корня. Придаточные корни могут возникать и на стеблях, и на листьях, и на корнях. По происхождению выделяют следующие типы корневых систем:
1) система главного корня представлена главным корнем (первого порядка) с боковыми корнями второго и последующих порядков (у многих кустарников и деревьев, большинства двудольных растений);
2) система придаточных корней развивается на стеблях, листьях; встречается у большинства однодольных растений и многих двудольных, размножающихся вегетативно;
3) смешанная корневая система образована главным и придаточными корнями с их боковыми ответвлениями (многие травянистые двудольные).
По форме различают стержневую и мочковатую корневые системы.
В стержневой корневой системе главный корень сильно развит и хорошо заметен среди остальных корней. В мочковатой корневой системе главный корень незаметен или его нет, а корневая система составлена многочисленными придаточными корнями.
Источник