
Ток воды по сосудам растений

- главная
- люди
- Петя Кантроп
Основные двигатели водного тока
Поглощение воды корневой системой идет благодаря работе двух концевых двигателей водного тока: верхнего концевого двигателя, или присасывающей силы испарения (транспирации), и нижнего концевого двигателя, или корневого двигателя. Основной силой, вызывающей поступление и передвижение воды в растении, является присасывающая сила транспирации, в результате которого возникает градиент водного потенциала. Водный потенциал – это мера энергии, используемой водой для передвижения. Водный потенциал и сосущая сила одинаковы по абсолютному значению, но противоположны по знаку. Чем меньше насыщенность водой данной системы, тем меньше (более отрицателен) ее водный потенциал. При потере воды растением в процессе транспирации создается ненасыщенность клеток листа водой, как следствие, возникает сосущая сила (водный потенциал падает). поступление воды идет в сторону большей сосущей силы, или меньшего водного потенциала.
Таким образом, верхний концевой двигатель водного тока в растении – это присасывающая сила транспирации листьев, и его работа мало связана с жизнедеятельностью корневой системы. Действительно, опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется.
Кроме верхнего концевого двигателя водного тока, в растениях существует нижний концевой двигатель. Это хорошо доказывается на примере таких явлениях, как гуттация.
Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, препятствующей испарению, выделяют капельно-жидкую воду с небольшим количеством растворенных веществ – гуттация. Выделение жидкости идет через специальные водные устьица – гидаторы. Выделяющаяся жидкость – гутта. Таким образом, процесс гуттации является результатом одностороннего тока воды, происходящего в отсутствие транспирации, и, следовательно, вызывается какой-то иной причиной.
К такому же выводу можно прийти и при рассмотрении явления плач растений. Если срезать побеги растения и к срезанному концу присоединить стеклянную трубку, то по ней будет подниматься жидкость. Анализ показывает, что это вода с растворенными веществами – пасока. В некоторых случаях, особенно в весенний период, плач наблюдается и при надрезе веток растений. Определения показали, что объем выделяющейся жидкости (пасоки) во много раз превышает объем корневой системы. Таким образом, плач – это не просто вытекание жидкости в результате пореза. Все сказанное приводит к выводу, что плач, как и гуттация, связана с наличием одностороннего тока воды через корневые системы, не зависящего от транспирации. Силу, вызывающую односторонний ток воды по сосудам с растворенными веществами, не зависящую от процесса транспирации, называют корневым давлением. Наличие корневого давления позволяет говорить о нижнем концевом двигателе водного тока. Корневое давление можно измерить, присоединив манометр к концу, оставшемуся после срезания надземных органов растения, или поместив корневую систему в серию растворов различной концентрации и подобрав такую, при которой плач прекращается. Оказалось, что корневое давление равняется примерно 0,1 – 0,15 МПа (Д.А.Сабинин). Определения, проведенные советскими исследователями Л.В.Можаевой, В.Н.Жолкевичем, показали, что концентрация наружного раствора, останавливающего плач, значительно выше концентрации пасоки. Это позволило высказать мнение, что плач может идти против градиента концентрации. Было показано также, что плач осуществляется только в тех условиях, в которых нормально протекают все процессы жизнедеятельности клеток. Не только умерщвление клеток корня, но и снижение интенсивности их жизнедеятельности, в первую очередь интенсивность дыхания, прекращает плач. В отсутствии кислорода, под влиянием дыхательных ядов, при понижении температуры плач приостанавливается. Все сказанное позволило Д.А.Сабинину дать следующее определение: плач растений – это прижизненный односторонний ток воды и питательных веществ, зависящий от аэробной переработки ассимелятов. Д.А.Сабинин предложил схему, объясняющую механизм одностороннего тока воды в корне. Согласно этой гипотезе, клетки корня поляризованы в определенном направлении. Это проявляется в том, что в разных отсеках одной и той же клетки процессы обмена веществ различны. В одной части клетки идут усиленные процессы распада, в частности, крахмала на сахара, вследствие чего концентрация клеточного сока возрастает. На противоположном конце клетки преобладают процессы синтеза, благодаря чему концентрация растворенных веществ в этой части клетки уменьшается. Надо учитывать, что все эти механизмы будут работать только при достаточном количестве воды в среде и не нарушенном обмене веществ.
Согласно другой гипотезе, зависимость плача растений от интенсивности дыхания является косвенной. Энергия дыхания используется для поступления ионов в клетки коры, откуда они десорбируются в сосуды ксилемы. В результате концентрация солей в сосудах ксилемы повышается, что и вызывает поступление воды.
Передвижение воды по растению
Вода, поглощенная клетками корня, под влиянием разности водных потенциалов, которые возникают благодаря транспирации, а также силе корневого давления, передвигается до проводящих путей ксилемы. Согласно современным представлениям, вода в корневой системе передвигается не только по живым клеткам. Еще в 1932 г. немецкий физиолог Мюнх развил представление о существовании в корневой системе двух относительно не зависимых друг от друга объемов, по которым передвигается вода, – апопласта и симпласта. Апопласт – это свободное пространство корня, в которое входят межклетные промежутки, оболочки клеток, а также сосуды ксилемы. Симпласт – это совокупность протопластов всех клеток, отграниченных полупроницаемой мембраной. Благодаря многочисленным плазмодесмам, соединяющим между собой протопласт отдельных клеток, симпласт представляет единую систему. Апопласт, по-видимому, не непрерывен, а разделен на два объема. Первая часть апопласта расположена в коре корня до клеток эндодермы, вторая – по другую сторону клеток эндодермы, и включает в себя сосуды ксилемы. Клетки эндодермы благодаря пояскам Каспари представляют как бы барьер для передвижения воды по свободному пространству (межклетникам и клеточным оболочкам). Для того чтоб попасть в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану и главным образом по апопласту и лишь частично по симпласту. Однако в клетках эндодермы передвижение воды идет, по-видимому, по симпласту. Далее вода поступает в сосуды ксилемы. Затем передвижение воды идет по сосудистой системе корня, стебля и листа.
Из сосудов стебля вода движется через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Иногда мелких ответвлений жилок листа так много, что они подводят воду почти к каждой клетке. Вся вода в клетке находится в равновесном состоянии. Иначе говоря, в смысле насыщенности водой, имеется равновесие между вакуолью, цитоплазмой и клеточной оболочкой, их водные потенциалы равны. Вода передвигается от клетки к клетке благодаря градиенту сосущей силы.
Вся вода в растении представляет единую взаимосвязанную систему. Поскольку между молекулами воды имеются силы сцепления (когезия), вода поднимается на высоту значительно большую 10 м. сила сцепления увеличивается, так как молекулы воды обладают большим сродством друг к другу. Силы сцепления обладают и между водой и стенками сосудов.
Степень натяжения водных нитей в сосудах зависит от соотношения процессов поглощения и испарения воды. Все это позволяет растительному организму поддерживать единую водную систему и не обязательно восполнять каждую каплю испаряемой воды.
В том случае, если в отдельные членики сосудов попадает воздух, они, по-видимому, выключается из общего тока проведения воды. Таков путь передвижения воды по растению (рис. 1).

Рис. 1. Путь воды в растении.
Скорость перемещения воды по растению в течение суток изменяется. В дневные часы она на много больше. При этом разные виды растений различаются по скорости передвижения воды. Изменение температуры, введение метаболических ингибиторов не влияют на передвижение воды. Вместе с тем этот процесс, как и следовало ожидать, очень сильно зависит от скорости транспирации и от диаметра водопроводящих сосудов. В более широких сосудах вода встречает меньшее сопротивление. Однако надо учитывать, что в более широкие сосуды могут попасть пузырьки воздуха или произойти какие-либо иные нарушения тока воды.
Видео: Движение воды и органических веществ по стеблю.
<iframe src=”https://www.youtube.com/embed/OfkoOseoD8E” frameborder=”0″ width=”420″ height=”315″></iframe>
Источник
- Главная
- Природа
- Проводящая система растений
Елена Голец30 Август 2017 6763
Проводящая система растений включает в себя такие понятия, как поступление, движение воды в растениях и испарение ее. Вода необходима растениям.
Вода необходима растениям.
Проводящая система растений
Передвигается вода в растениях по клеткам коровой паренхимы до центрального цилиндра корня, затем по проводящей системе до листовой паренхимы и, наконец, по клеткам листовой паренхимы. На первом участке пути вода передвигается осмотически благодаря повышению сосущей силы клеток корня. Движение воды от корневого волоска в сторону центральных сосудов.
Этот отрезок пути очень небольшой (доли миллиметра), но передвижение воды по этому участку очень затруднено, так как воде приходится преодолевать сопротивление слоев живой протоплазмы. Это сопротивление примерно равно 1 атм на 1 мм пути, поэтому передвижение воды по живым клеткам на более значительные расстояния не обеспечивало бы потребности растения в воде.
Действительно, растения, у которых не развита проводящая система, например мхи, (подробнее: Как образуется болото) имеют незначительные размеры и приспособлены к жизни только во влажных условиях. У наземных растений в процессе эволюции образовалась проводящая ткань, которая устанавливает сообщение между всасывающими воду корнями и испаряющими воду листьями.
Проводящая воду ткань
Проводящая воду ткань состоит из сосудов, или трахей, и трахеидов; она начинается в центральном цилиндре корня, проходит через весь корень и стебель и заканчивается в виде тончайших разветвлений — жилок, пронизывающих всю листовую паренхиму.
Сосуды представляют собой мертвые трубки, образовавшиеся из живых клеток. В сосудах сохраняются поперечные перегородки на разном расстоянии (от нескольких миллиметров до метра в зависимости от вида растения) одна от другой.
Исчезновение перегородок даже на небольшом расстоянии в тысячи раз ускоряет передвижение воды. Трахеиды это длинные мертвые клетки с заостренными концами. При образовании сосудов и трахеид происходит утолщение и одревеснение их оболочек, вследствие чего они не сдавливаются под давлением окружающих их живых паренхимных клеток.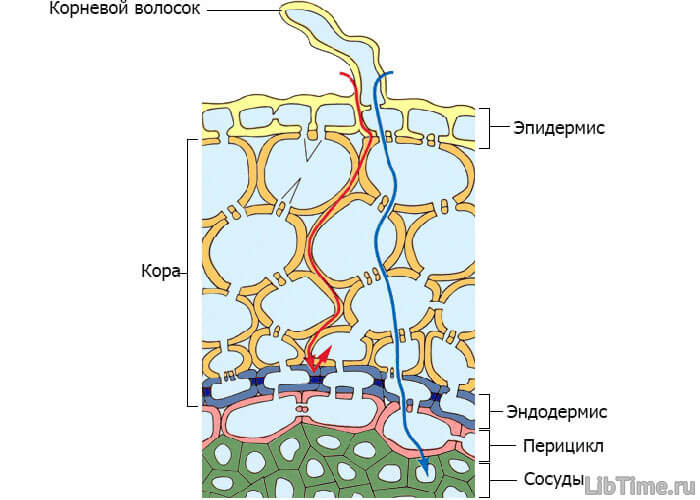 Движение воды в древесных растениях.
Движение воды в древесных растениях.
Одревеснение, однако, никогда не бывает сплошным: на стенке сосудов остаются тонкие места — поры, по которым вода может перемещаться не только вверх по сосудам, но и в радиальном направлении.
Подъем воды по сосудам
Подъем воды по сосудам можно доказать следующим опытом. Если у срезанной и поставленной в воду ветки снять кольцо коры выше уровня воды, листья ее не завянут, так как сосуды расположены в древесине.
Движение воды по сосудам чаще всего направлено снизу вверх и называется поэтому восходящим током.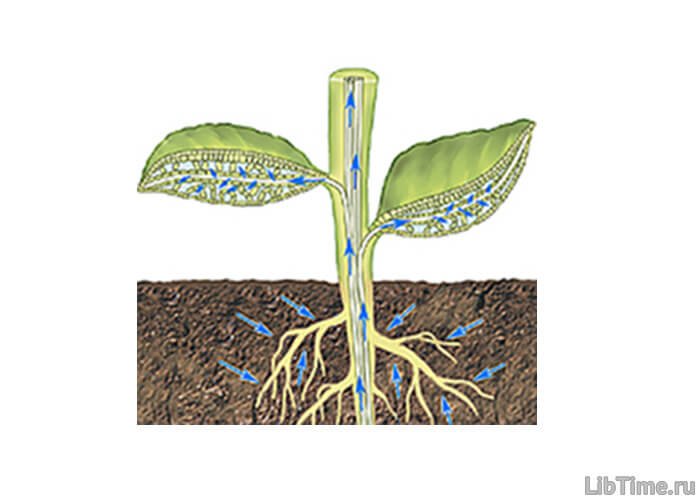 Восходящий ток.
Восходящий ток.
Последний отрезок пути водного тока по листовой паренхиме идет по живым
клеткам. Вода передвигается осмотическим путем по клеткам мезофилла листа до последних клеток, граничащих с межклеточниками. Этот отрезок пути, так же как и первый, очень короткий.
Если срезанную ветку растения герметически закрепить в стеклянной трубке, заполненной водой, и нижний конец ее опустить в сосуд со ртутью, то при испарении веткой воды ртуть в трубке будет подниматься.
Из этого опыта ясно, что передвижение воды по растению обусловлено главным образом транспирацией, (подробнее: Транспирация у растений), а не только корневым давлением.
При испарении воды с поверхности листьев в клетках возникает сосущая сила. Величина ее тем больше, чем меньше воды остается в клетках листа. Эта возникающая сосущая сила поддерживает постоянное передвижение воды в растении.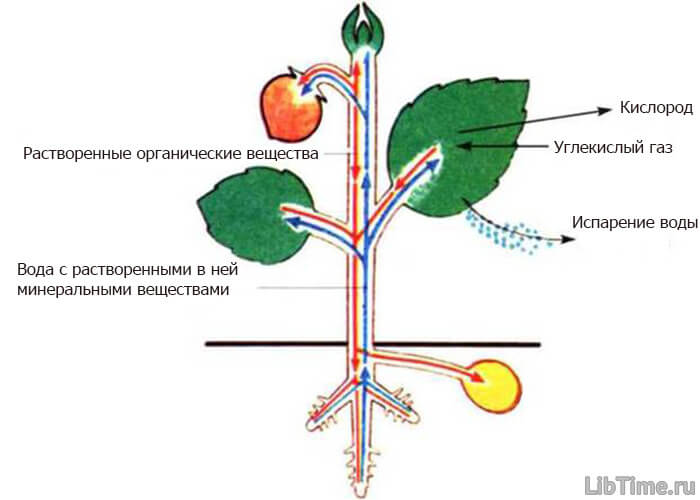 Транспорт веществ в растениях.
Транспорт веществ в растениях.
Силы, приводящие воду в движение
Таким образом, силы, приводящие воду в движение, находятся по концам проводящей системы: нагнетающий воду корень, работа которого получила название нижнего концевого двигателя, и сила присасывания воды листьями — верхний концевой двигатель.
Оба двигателя действуют в одном направлении и могут заменять и дополнять друг друга. Во время сильной инсоляции летом и при, засухе водоснабжение растения идет за счет присасывающего действия транспирации.
Корневое давление
Когда же почва богата водой, а воздух водяными парами, подъем воды обеспечивается силой корневого давления, (подробнее: Корни обеспечивают растение водой). Следовательно, в зависимости от условий внешней среды главная роль принадлежит то одному, то другому концевому двигателю.
Водные нити не рвутся под влиянием своей тяжести, несмотря на то, что при сильной транспирации они находятся в состоянии натяжения. Это объясняется силой сцепления молекул воды, достигающей 300—350 атм, а так как в сосудах нет воздуха, то целостность водного тока не прерывается.
Скорость водного тока
Скорость водного тока зависит от строения проводящих воду элементов. Вода быстрее передвигается по сосудам, причем скорость движения ее зависит от диаметра сосудов: чем он меньше, тем медленнее будет передвигаться вода.
Движение воды в растениях происходит благодаря работе двух концевых двигателей, верхнего и нижнего, и сил сцепления, обеспечивающих целостность водных нитей.
Рейтинг: 3/5 – 1
голосов
Источник
ÐÑновной Ñилой, вÑзÑваÑÑей поÑÑÑпление и пеÑедвижение Ð²Ð¾Ð´Ñ Ð² ÑаÑÑении, ÑвлÑеÑÑÑ Ð¿ÑоÑеÑÑ ÑÑанÑпиÑаÑии, в ÑезÑлÑÑаÑе коÑоÑого Ð²Ð¾Ð·Ð½Ð¸ÐºÐ°ÐµÑ Ð³ÑÐ°Ð´Ð¸ÐµÐ½Ñ Ð²Ð¾Ð´Ð½Ð¾Ð³Ð¾ поÑенÑиала. Ðак Ñже ÑпоминалоÑÑ, воднÑй поÑенÑиал â ÑÑо меÑа ÑнеÑгии, иÑполÑзÑемой водой Ð´Ð»Ñ Ð¿ÐµÑедвижениÑ. Чем менÑÑе наÑÑÑенноÑÑÑ Ð²Ð¾Ð´Ð¾Ð¹ данной ÑиÑÑемÑ, Ñем менÑÑе (более оÑÑиÑаÑелен) ее воднÑй поÑенÑиал. ÐÑи поÑеÑе Ð²Ð¾Ð´Ñ ÑаÑÑением в пÑоÑеÑÑе ÑÑанÑпиÑаÑии ÑоздаеÑÑÑ Ð½ÐµÐ½Ð°ÑÑÑенноÑÑÑ ÐºÐ»ÐµÑок лиÑÑа водой, как ÑледÑÑвие, воднÑй поÑенÑиал ÑÑановиÑÑÑ Ð±Ð¾Ð»ÐµÐµ оÑÑиÑаÑелÑнÑм. ÐоÑÑÑпление Ð²Ð¾Ð´Ñ Ð¸Ð´ÐµÑ Ð² ÑÑоÑÐ¾Ð½Ñ Ð±Ð¾Ð»ÐµÐµ оÑÑиÑаÑелÑного водного поÑенÑиала.
Так назÑваемÑй веÑÑ Ð½Ð¸Ð¹ конÑевой двигаÑÐµÐ»Ñ Ð²Ð¾Ð´Ð½Ð¾Ð³Ð¾ Ñока в ÑаÑÑении â ÑÑо ÑÑанÑпиÑаÑÐ¸Ñ Ð»Ð¸ÑÑÑев. Ðднако опÑÑÑ Ð¿Ð¾ÐºÐ°Ð·Ð°Ð»Ð¸, ÑÑо вода Ð¼Ð¾Ð¶ÐµÑ Ð¿Ð¾ÑÑÑпаÑÑ Ð² побеги и ÑеÑез меÑÑвÑÑ ÐºÐ¾ÑневÑÑ ÑиÑÑемÑ, пÑиÑем в ÑÑом ÑлÑÑае поглоÑение Ð²Ð¾Ð´Ñ Ð´Ð°Ð¶Ðµ ÑÑкоÑÑеÑÑÑ. ÐÑоме веÑÑ Ð½ÐµÐ³Ð¾ конÑевого двигаÑÐµÐ»Ñ Ð²Ð¾Ð´Ð½Ð¾Ð³Ð¾ Ñока, в ÑаÑÑениÑÑ ÑÑÑеÑÑвÑÐµÑ Ð½Ð¸Ð¶Ð½Ð¸Ð¹ конÑевой двигаÑелÑ. ÐÑо Ñ Ð¾ÑоÑо доказÑваеÑÑÑ Ð½Ð° пÑимеÑе ÑÐ°ÐºÐ¸Ñ Ñвлений, как гÑÑÑаÑÐ¸Ñ Ð¸ плаÑ. ÐиÑÑÑÑ ÑаÑÑений, клеÑки коÑоÑÑÑ Ð½Ð°ÑÑÑÐµÐ½Ñ Ð²Ð¾Ð´Ð¾Ð¹, в ÑÑловиÑÑ Ð²ÑÑокой влажноÑÑи воздÑÑ Ð°, пÑепÑÑÑÑвÑÑÑей иÑпаÑениÑ, вÑделÑÑÑ ÐºÐ°Ð¿ÐµÐ»Ñно-жидкÑÑ Ð²Ð¾Ð´Ñ Ñ Ð½ÐµÐ±Ð¾Ð»ÑÑим колиÑеÑÑвом ÑаÑÑвоÑеннÑÑ Ð²ÐµÑеÑÑв â гÑÑÑиÑÑÑÑ. ÐÑделение жидкоÑÑи Ð¸Ð´ÐµÑ ÑеÑез ÑпеÑиалÑнÑе воднÑе ÑÑÑÑиÑа â гидаÑодÑ. ÐÑделÑÑÑаÑÑÑ Ð¶Ð¸Ð´ÐºÐ¾ÑÑÑ â гÑÑÑа. Таким обÑазом, пÑоÑеÑÑ Ð³ÑÑÑаÑии ÑвлÑеÑÑÑ ÑезÑлÑÑаÑом одноÑÑоÑоннего Ñока водÑ, пÑоиÑÑ Ð¾Ð´ÑÑего в оÑÑÑÑÑÑвие ÑÑанÑпиÑаÑии, и, ÑледоваÑелÑно, вÑзÑваеÑÑÑ ÐºÐ°ÐºÐ¾Ð¹-Ñо иной пÑиÑиной. Ð ÑÐ°ÐºÐ¾Ð¼Ñ Ð¶Ðµ вÑÐ²Ð¾Ð´Ñ Ð¼Ð¾Ð¶Ð½Ð¾ пÑийÑи и пÑи ÑаÑÑмоÑÑении ÑÐ²Ð»ÐµÐ½Ð¸Ñ Ð¿Ð»Ð°Ñа ÑаÑÑений. ÐÑли ÑÑезаÑÑ Ð¿Ð¾Ð±ÐµÐ³Ð¸ ÑаÑÑÐµÐ½Ð¸Ñ Ð¸ к ÑÑÐµÐ·Ð°Ð½Ð½Ð¾Ð¼Ñ ÐºÐ¾Ð½ÑÑ Ð¿ÑиÑоединиÑÑ ÑÑеклÑннÑÑ ÑÑÑбкÑ, Ñо по ней бÑÐ´ÐµÑ Ð¿Ð¾Ð´Ð½Ð¸Ð¼Ð°ÑÑÑÑ Ð¶Ð¸Ð´ÐºÐ¾ÑÑÑ. Ðнализ показÑваеÑ, ÑÑо ÑÑо â вода Ñ ÑаÑÑвоÑеннÑми веÑеÑÑвами, полÑÑивÑÐ°Ñ Ð½Ð°Ð·Ð²Ð°Ð½Ð¸Ðµ паÑоки. РнекоÑоÑÑÑ ÑлÑÑаÑÑ , оÑобенно в веÑенний пеÑиод, Ð¿Ð»Ð°Ñ Ð½Ð°Ð±Ð»ÑдаеÑÑÑ Ð¸ пÑи надÑезе веÑок ÑаÑÑений. Ðменно на ÑÑом оÑновано вÑÑекание Ñока пÑи надÑезе ÑÑвола беÑÐµÐ·Ñ Ð²ÐµÑной. ÐпÑÐµÐ´ÐµÐ»ÐµÐ½Ð¸Ñ Ð¿Ð¾ÐºÐ°Ð·Ð°Ð»Ð¸, ÑÑо обÑем вÑделÑÑÑейÑÑ Ð¶Ð¸Ð´ÐºÐ¾ÑÑи (паÑоки) во много Ñаз пÑевÑÑÐ°ÐµÑ Ð¾Ð±Ñем коÑневой ÑиÑÑемÑ. Таким обÑазом, Ð¿Ð»Ð°Ñ â ÑÑо не пÑоÑÑо вÑÑекание жидкоÑÑи в ÑезÑлÑÑаÑе поÑеза. ÐÑо пÑÐ¸Ð²Ð¾Ð´Ð¸Ñ Ðº вÑводÑ, ÑÑо плаÑ, как и гÑÑÑаÑиÑ, ÑвÑзан Ñ Ð½Ð°Ð»Ð¸Ñием одноÑÑоÑоннего Ñока Ð²Ð¾Ð´Ñ ÑеÑез коÑневÑе ÑиÑÑемÑ, не завиÑÑÑего Ð¾Ñ ÑÑанÑпиÑаÑии.
СилÑ, вÑзÑваÑÑÑÑ Ð² ÑаÑÑении одноÑÑоÑонний Ñок Ð²Ð¾Ð´Ñ Ñ ÑаÑÑвоÑеннÑми веÑеÑÑвами, не завиÑÑÑÑÑ Ð¾Ñ Ð¿ÑоÑеÑÑа ÑÑанÑпиÑаÑии, назÑваÑÑ ÐºÐ¾ÑневÑм давлением. ÐалиÑие коÑневого Ð´Ð°Ð²Ð»ÐµÐ½Ð¸Ñ Ð¿Ð¾Ð·Ð²Ð¾Ð»ÑÐµÑ Ð³Ð¾Ð²Ð¾ÑиÑÑ Ð¾ нижнем конÑевом двигаÑеле водного Ñока. ÐоÑневое давление можно измеÑиÑÑ, пÑиÑоединив маномеÑÑ Ðº конÑÑ, оÑÑавÑемÑÑÑ Ð¿Ð¾Ñле ÑÑÐµÐ·Ð°Ð½Ð¸Ñ Ð½Ð°Ð´Ð·ÐµÐ¼Ð½ÑÑ Ð¾Ñганов ÑаÑÑениÑ, или помеÑÑив коÑневÑÑ ÑиÑÑÐµÐ¼Ñ Ð² ÑеÑÐ¸Ñ ÑаÑÑвоÑов ÑазлиÑной конÑенÑÑаÑии и подобÑав ÑакÑÑ, пÑи коÑоÑой Ð¿Ð»Ð°Ñ Ð¿ÑекÑаÑаеÑÑÑ. ÐказалоÑÑ, ÑÑо велиÑина коÑневого Ð´Ð°Ð²Ð»ÐµÐ½Ð¸Ñ Ñавна пÑимеÑно 1â1,5 баÑа (ÐÐ. Сабинин). ÐÑло показано Ñакже, ÑÑо Ð¿Ð»Ð°Ñ Ð¾ÑÑÑеÑÑвлÑеÑÑÑ ÑолÑко в ÑÐµÑ ÑÑловиÑÑ , в коÑоÑÑÑ Ð½Ð¾ÑмалÑно пÑоÑекаÑÑ Ð²Ñе пÑоÑеÑÑÑ Ð¶Ð¸Ð·Ð½ÐµÐ´ÐµÑÑелÑноÑÑи клеÑок коÑнÑ. Ðе ÑолÑко ÑмеÑÑвление клеÑок коÑнÑ, но и Ñнижение инÑенÑивноÑÑи Ð¸Ñ Ð¶Ð¸Ð·Ð½ÐµÐ´ÐµÑÑелÑноÑÑи, в пеÑвÑÑ Ð¾ÑеÑÐµÐ´Ñ Ð¸Ð½ÑенÑивноÑÑи дÑÑ Ð°Ð½Ð¸Ñ, пÑекÑаÑÐ°ÐµÑ Ð¿Ð»Ð°Ñ. РоÑÑÑÑÑÑвие киÑлоÑода, под влиÑнием дÑÑ Ð°ÑелÑнÑÑ Ñдов, пÑи пониженной ÑемпеÑаÑÑÑе Ð¿Ð»Ð°Ñ Ð¿ÑиоÑÑанавливаеÑÑÑ. ÐÑе Ñказанное позволило Ð. Ð. Ð¡Ð°Ð±Ð¸Ð½Ð¸Ð½Ñ Ð´Ð°ÑÑ ÑледÑÑÑее опÑеделение: Ð¿Ð»Ð°Ñ ÑаÑÑений â ÑÑо пÑижизненнÑй одноÑÑоÑонний Ñок Ð²Ð¾Ð´Ñ Ð¸ пиÑаÑелÑнÑÑ Ð²ÐµÑеÑÑв, ÑвÑзаннÑй Ñ Ð·Ð°ÑÑаÑой ÑнеÑгии. Ðднако Ð¼ÐµÑ Ð°Ð½Ð¸Ð·Ð¼ ÑÑого ÑÐ²Ð»ÐµÐ½Ð¸Ñ Ð´Ð¾ наÑÑоÑÑего вÑемени не полÑÑил доÑÑаÑоÑного обÑÑÑнениÑ. СÑÑеÑÑвÑÑÑ Ð´Ð²Ðµ оÑновнÑе ÑоÑки зÑениÑ. Ð Ñд иÑÑледоваÑелей (Ð. ÐÑаÑÑÑ, Т. ÐÑÐ¾Ð¹ÐµÑ Ð¸ дÑ.) полагаÑÑ, ÑÑо вода пеÑедвигаеÑÑÑ Ð¿Ð°ÑÑивно, оÑмоÑиÑеÑким пÑÑем по гÑадиенÑÑ Ð²Ð¾Ð´Ð½Ð¾Ð³Ð¾ поÑенÑиала. СоглаÑно ÑÑой гипоÑезе низкий (более оÑÑиÑаÑелÑнÑй) воднÑй поÑенÑиал ÑоздаеÑÑÑ Ð² ÑоÑÑÐ´Ð°Ñ ÐºÑÐ¸Ð»ÐµÐ¼Ñ Ð±Ð»Ð°Ð³Ð¾Ð´Ð°ÑÑ Ð¿Ð¾ÑÑÑÐ¿Ð»ÐµÐ½Ð¸Ñ ÑÑда из поÑÐ²Ñ ÑаÑÑвоÑимÑÑ Ñолей, а Ñакже из-за Ñлабого пÑоÑÐ¸Ð²Ð¾Ð´Ð°Ð²Ð»ÐµÐ½Ð¸Ñ Ð¼Ð°Ð»Ð¾ÑлаÑÑиÑнÑÑ ÑÑенок ÑоÑÑдов. Соли поÑÑÑпаÑÑ Ð°ÐºÑивно, Ð´Ð»Ñ Ñего и ÑÑебÑеÑÑÑ Ð·Ð°ÑÑаÑа ÑнеÑгии. Таким обÑазом, поÑÑÑпление Ð²Ð¾Ð´Ñ ÑвÑзано в ÑÑом ÑлÑÑае Ñ ÑнеÑгеÑиÑеÑкими заÑÑаÑами и Ñ Ð°ÐºÑивной жизнедеÑÑелÑноÑÑÑÑ ÐºÐ»ÐµÑок коÑÐ½Ñ Ð½Ðµ пÑÑмо, а коÑвенно. Ð ÑÑой ÑвÑзи коÑневое давление можно опÑеделиÑÑ ÐºÐ°Ðº давление, ÑазвиваÑÑееÑÑ Ð² пÑоводÑÑÐ¸Ñ ÑлеменÑÐ°Ñ ÐºÑÐ¸Ð»ÐµÐ¼Ñ Ð±Ð»Ð°Ð³Ð¾Ð´Ð°ÑÑ Ð¼ÐµÑаболиÑеÑкой акÑивноÑÑи коÑней. ÐоÑколÑÐºÑ Ð°ÐºÑивное поÑÑÑпление завиÑÐ¸Ñ Ð¾Ñ ÑнеÑгии, вÑделÑемой пÑи дÑÑ Ð°Ð½Ð¸Ð¸, ÑабоÑа нижнего конÑевого двигаÑÐµÐ»Ñ â коÑневого Ð´Ð°Ð²Ð»ÐµÐ½Ð¸Ñ Ð·Ð°Ð²Ð¸ÑÐ¸Ñ Ð¾Ñ Ð¿ÑиÑÑÑÑÑÐ²Ð¸Ñ ÐºÐ¸ÑлоÑода, ингибиÑоÑов и акÑиваÑоÑов дÑÑ Ð°ÑелÑного обмена.
ÐмеÑÑе Ñ Ñем ÑÑÑеÑÑвÑÐµÑ Ð¸ дÑÑÐ³Ð°Ñ ÑоÑка зÑÐµÐ½Ð¸Ñ (Ð.Ð. ÐолкевиÑ, Ð.Ð. Ðожаева, Ð.Ð. ÐилÑÑиков), ÑоглаÑно коÑоÑой коÑневое давление ÑкладÑваеÑÑÑ Ð¸Ð· двÑÑ ÑоÑÑавлÑÑÑÐ¸Ñ : оÑмоÑиÑеÑкой и меÑаболиÑеÑкой. РабоÑа поÑледней ÑÑебÑÐµÑ Ð½ÐµÐ¿Ð¾ÑÑедÑÑвенной заÑÑаÑÑ ÑнеÑгии ÐТФ. ÐÑÑказÑваеÑÑÑ Ð¿Ñедположение, ÑÑо пÑи ÑÑом болÑÑÐ°Ñ ÑÐ¾Ð»Ñ Ð¿ÑÐ¸Ð½Ð°Ð´Ð»ÐµÐ¶Ð¸Ñ ÑокÑаÑиÑелÑнÑм акÑиноподобнÑм белкам, ÑнеÑгозавиÑимое ÑокÑаÑение и ÑаÑÑлабление коÑоÑÑÑ Ð²ÑзÑÐ²Ð°ÐµÑ Ð¸Ð·Ð¼ÐµÐ½ÐµÐ½Ð¸Ñ Ð³Ð¸Ð´ÑоÑÑаÑиÑеÑкого Ð´Ð°Ð²Ð»ÐµÐ½Ð¸Ñ Ð² клеÑÐºÐ°Ñ . Ð ÑезÑлÑÑаÑе на пÑÑи водного Ñока в напÑавлении ÑоÑÑдов кÑÐ¸Ð»ÐµÐ¼Ñ ÑоздаÑÑÑÑ Ð»Ð¾ÐºÐ°Ð»ÑнÑе гÑадиенÑÑ Ð²Ð¾Ð´Ð½Ð¾Ð³Ð¾ поÑенÑиала, ÑÑо и ÑпоÑобÑÑвÑÐµÑ Ð¿ÑоÑÐ°Ð»ÐºÐ¸Ð²Ð°Ð½Ð¸Ñ Ð²Ð¾Ð´Ñ Ð² ÑоÑÑдÑ. ÐопÑÐ¾Ñ ÑÑÐ¾Ñ ÑÑебÑÐµÑ Ð´Ð°Ð»ÑнейÑей ÑкÑпеÑименÑалÑной ÑазÑабоÑки. Сила, ÑÐ°Ð·Ð²Ð¸Ð²Ð°ÐµÐ¼Ð°Ñ Ð½Ð¸Ð¶Ð½Ð¸Ð¼ конÑевÑм двигаÑелем водного Ñока, во много Ñаз менÑÑе Ñой, коÑоÑÐ°Ñ Ð¾Ð±ÑÑловливаеÑÑÑ ÑÑанÑпиÑаÑией. ÐÑо пÑоÑвлÑеÑÑÑ Ð¸ в Ñом, ÑÑо ÑкоÑоÑÑÑ Ð²ÑÑÐµÐºÐ°Ð½Ð¸Ñ Ð²Ð¾Ð´Ñ Ð¸Ð· кÑÐ¸Ð»ÐµÐ¼Ñ Ð·Ð½Ð°ÑиÑелÑно ÑÑÑÑÐ¿Ð°ÐµÑ ÑкоÑоÑÑи ÑÑанÑпиÑаÑии, поÑÑÐ¾Ð¼Ñ Ð² обÑÑнÑÑ ÑÑловиÑÑ Ð²Ð¾Ð´Ð° в ÑоÑÑÐ´Ð°Ñ Ð½Ð°Ñ Ð¾Ð´Ð¸ÑÑÑ Ð½Ðµ под положиÑелÑнÑм, а под оÑÑиÑаÑелÑнÑм давлением. Ðднако вÑе же нижний конÑевой двигаÑÐµÐ»Ñ ÑпоÑобÑÑвÑÐµÑ Ð¿Ð¾Ð´Ð°Ñе водÑ. ÐолÑÑинÑÑво иÑÑледоваÑелей ÑÑиÑаеÑ, ÑÑо пÑекÑаÑение ÑабоÑÑ Ð½Ð¸Ð¶Ð½ÐµÐ³Ð¾ конÑевого двигаÑÐµÐ»Ñ ÑвлÑеÑÑÑ Ð¿Ð¾ÐºÐ°Ð·Ð°Ñелем наÑала ÑÑÑÐ°Ð´Ð°Ð½Ð¸Ñ ÑаÑÑений Ð¾Ñ Ð½ÐµÐ´Ð¾ÑÑаÑка Ð²Ð¾Ð´Ñ (заÑÑÑ Ð¸). ÐÑобенное знаÑение ÑÑÐ¾Ñ Ð´Ð²Ð¸Ð³Ð°ÑÐµÐ»Ñ Ð²Ð¾Ð´Ð½Ð¾Ð³Ð¾ Ñока пÑиобÑеÑÐ°ÐµÑ Ð² оÑÑÑÑÑÑвие ÑÑанÑпиÑаÑии. Так, ÑÑо Ð¼Ð¾Ð¶ÐµÑ Ð¸Ð¼ÐµÑÑ Ð¼ÐµÑÑо Ñанней веÑной, когда воздÑÑ Ð½Ð°ÑÑÑен водой и ÑÑанÑпиÑаÑÐ¸Ñ Ð¾Ñлаблена. У некоÑоÑÑÑ Ð¼Ð½Ð¾Ð³Ð¾Ð»ÐµÑÐ½Ð¸Ñ ÑаÑÑений в зимний пеÑиод ÑоÑÑÐ´Ñ ÐºÑÐ¸Ð»ÐµÐ¼Ñ Ð·Ð°Ð¿Ð¾Ð»Ð½ÐµÐ½Ñ Ð²Ð¾Ð·Ð´ÑÑ Ð¾Ð¼ и веÑной поÑÑÑпление Ð²Ð¾Ð´Ñ Ð¸Ð´ÐµÑ Ð·Ð° ÑÑÐµÑ ÑабоÑÑ ÑолÑко нижнего конÑевого двигаÑелÑ.
Источник